Les homininés regroupent l’ensemble des espèces apparentées à l’être humain actuel (Homo sapiens) après leur séparation avec la lignée des chimpanzés. Les analyses paléogénétiques et les fossiles suggèrent que cette divergence s’est produite entre 7 et 6 millions d’années avant notre ère, dans un contexte de transformations climatiques majeures en Afrique.
Le premier représentant reconnu est Sahelanthropus tchadensis (≈ 7 Ma), découvert au Tchad. Son crâne, doté d’un foramen magnum avancé, témoigne d’une posture partiellement bipède. Il est suivi par Orrorin tugenensis (≈ 6 Ma, Kenya) et Ardipithecus kadabba (5,8 – 5,2 Ma), qui présentent déjà des adaptations locomotrices indiquant la transition d’un mode de vie arboricole vers la bipédie.
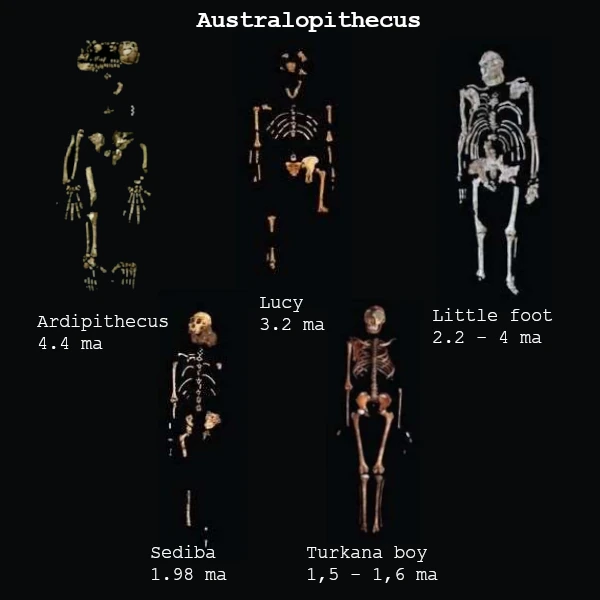
Entre 4,5 et 2 Ma, les australopithèques s’imposent comme les homininés dominants. Australopithecus afarensis (3,9 – 3,0 Ma), connu par le célèbre fossile « Lucy », incarne une bipédie plus affirmée, bien que leur capacité crânienne (~400 cm³) reste proche de celle des grands singes. Ces espèces sont contemporaines d’une diversification écologique, occupant des niches variées, allant de milieux boisés à des savanes plus ouvertes.
Vers 2,5 Ma, l’apparition du genre Homo marque un tournant évolutif majeur. Homo habilis et Homo rudolfensis sont associés aux premiers outils lithiques (industrie oldowayenne), témoignant de capacités cognitives nouvelles. Rapidement, Homo erectus (1,9 Ma) devient le premier grand colonisateur du monde, quittant l’Afrique pour peupler l’Asie et l’Europe, maîtrisant le feu et développant des sociétés organisées.
Au cours du dernier million d’années, la lignée humaine se complexifie avec Homo heidelbergensis, Homo neanderthalensis, les mystérieux Denisoviens, et des espèces insulaires telles que Homo floresiensis et Homo luzonensis. Enfin, Homo sapiens, apparu en Afrique il y a environ 300 000 ans, se distingue par l’émergence du langage symbolique, de l’art et de la culture cumulative, aboutissant à la diversité et à la domination planétaire actuelles.
Ainsi, l’évolution des homininés, s’étendant sur plus de 7 millions d’années, illustre un processus complexe de diversification et d’extinction, où plusieurs espèces ont cohabité avant que notre espèce ne devienne la seule survivante du genre Homo. Cette évolution buissonnante démontre que l’humanité n’est pas le résultat d’une progression linéaire, mais d’un enchevêtrement de lignées parallèles façonnées par la sélection naturelle, le climat et la culture.
| Espèce | Période (Ma) | Lieu principal | Caractéristiques clés |
|---|---|---|---|
| Sahelanthropus tchadensis | ~7,0 | Tchad | Crâne aplati, position du foramen magnum suggérant bipédie partielle |
| Orrorin tugenensis | 6,0 | Kenya | Fémur adapté à la bipédie, dents primitives |
| Ardipithecus kadabba | 5,8 – 5,2 | Éthiopie | Transition entre primates arboricoles et bipèdes |
| Ardipithecus ramidus | 4,4 | Éthiopie | Mixte arboricole et bipède, mains préhensiles |
| Australopithecus anamensis | 4,2 – 3,9 | Kenya | Bipédie affirmée, dentition primitive |
| Australopithecus afarensis | 3,9 – 3,0 | Éthiopie, Tanzanie | Lucy ; bipédie efficace, volume crânien ~400 cm³ |
| Australopithecus prometheus (Little Foot) | 3,67 | Afrique du Sud (Sterkfontein) | Squelette presque complet, bipédie et vie arboricole |
| Australopithecus bahrelghazali | 3,5 | Tchad | Découvert loin de la vallée du Rift, adaptation diverse |
| Australopithecus africanus | 3,0 – 2,1 | Afrique du Sud | Crâne de l’enfant de Taung, bipédie développée |
| Australopithecus garhi | 2,5 | Éthiopie | Associé aux premiers outils lithiques |
| Australopithecus sediba | 2,0 | Afrique du Sud | Traits intermédiaires entre australopithèques et Homo |
| Paranthropus aethiopicus | 2,7 – 2,3 | Kenya | Crâne noir, crête sagittale marquée |
| Paranthropus boisei | 2,3 – 1,2 | Afrique de l’Est | Mâchoires massives, régime végétarien spécialisé |
| Paranthropus robustus | 2,0 – 1,5 | Afrique du Sud | Dents très larges, adaptation au broyage |
| Homo habilis | 2,4 – 1,6 | Afrique de l’Est | Premier utilisateur d’outils Oldowayens |
| Homo rudolfensis | 2,4 – 1,8 | Kenya | Volume crânien plus grand que H. habilis |
| Homo erectus | 1,9 – 0,1 | Afrique, Asie, Europe | Premier grand migrant hors d’Afrique, maîtrise du feu |
| Homo antecessor | 1,2 – 0,8 | Espagne | Possible ancêtre commun de Néandertaliens et Sapiens |
| Homo heidelbergensis | 0,7 – 0,2 | Europe, Afrique | Chasseur organisé, ancêtre probable de H. sapiens et H. neanderthalensis |
| Homo floresiensis | 0,1 – 0,06 | Indonésie (Florès) | « Hobbit », petite taille (~1 m), cerveau réduit |
| Homo naledi | ~0,3 | Afrique du Sud | Comportement rituel funéraire probable |
| Homo luzonensis | 0,07 | Philippines | Espèce insulaire, mélange de traits archaïques |
| Homo neanderthalensis | 0,4 – 0,04 | Europe, Asie | Culture complexe, langage articulé probable |
| Homo longi | 0,3 – 0,15 | Chine | « Homme dragon », proche de Néandertal et Denisoviens |
| Homo sapiens | 0,3 – aujourd’hui | Afrique puis monde entier | Langage symbolique, pensée abstraite, culture cumulative |
Sources: Smithsonian Human Origins Program, MNHN Paris, Leakey Foundation, Berger et al. (2015), Stringer (2022).
 Déséquilibre Fatal: La Puissance Insoupçonnée de l'Infiniment Petit
Déséquilibre Fatal: La Puissance Insoupçonnée de l'Infiniment Petit  Le Grand Filtre de l'Évolution: la clé du paradoxe de Fermi
Le Grand Filtre de l'Évolution: la clé du paradoxe de Fermi  Pourquoi le tournesol se tourne-t-il vers le soleil? Une réponse par le Lagrangien
Pourquoi le tournesol se tourne-t-il vers le soleil? Une réponse par le Lagrangien  Population mondiale 2026: tendances démographiques par continent
Population mondiale 2026: tendances démographiques par continent  Pourquoi la vie émerge du déséquilibre et disparaît à l'équilibre thermodynamique
Pourquoi la vie émerge du déséquilibre et disparaît à l'équilibre thermodynamique  Le Spectre Électromagnétique et la Vision: Ce que nos yeux perçoivent de notre planète
Le Spectre Électromagnétique et la Vision: Ce que nos yeux perçoivent de notre planète  Le Soi et le Non-Soi: Une Lecture Physique Simplifiée de l'Identité
Le Soi et le Non-Soi: Une Lecture Physique Simplifiée de l'Identité  L’Horloge Moléculaire: Du hasard des mutations à la mesure du temps
L’Horloge Moléculaire: Du hasard des mutations à la mesure du temps  Les Empreintes de White Sands: Premiers Pas d’Amérique
Les Empreintes de White Sands: Premiers Pas d’Amérique 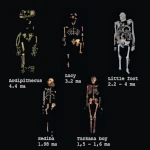 Homininés: Apparition, Expansion et Extinctions
Homininés: Apparition, Expansion et Extinctions  Catastrophes Naturelles Majeures: Quelles Sont les Menaces les Plus Probables?
Catastrophes Naturelles Majeures: Quelles Sont les Menaces les Plus Probables?  Les Grands Effondrements Civilisationnels: Périodes clés et Causes
Les Grands Effondrements Civilisationnels: Périodes clés et Causes 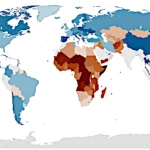 Naissances en Berne: Catastrophe Démographique ou Évolution Naturelle?
Naissances en Berne: Catastrophe Démographique ou Évolution Naturelle?  Sélection naturelle vs. Hasard: Pourquoi l’évolution n’est pas une loterie?
Sélection naturelle vs. Hasard: Pourquoi l’évolution n’est pas une loterie?  Et si la Vie Partait de la Terre? Une Révolution dans la Théorie de la Panspermie
Et si la Vie Partait de la Terre? Une Révolution dans la Théorie de la Panspermie  La Grande Bifurcation qui va Bouleverser Notre Monde: Survie ou Effondrement?
La Grande Bifurcation qui va Bouleverser Notre Monde: Survie ou Effondrement?  Chimie primordiale: Où naissent les premières molécules organiques?
Chimie primordiale: Où naissent les premières molécules organiques?  CO et CO₂: Deux Gaz, Deux Risques, Deux Mécanismes Biologiques
CO et CO₂: Deux Gaz, Deux Risques, Deux Mécanismes Biologiques  Synchronisation Spontanée: un Phénomène Universel, de la Physique au Vivant
Synchronisation Spontanée: un Phénomène Universel, de la Physique au Vivant  Défi temporel: comment illustrer un milliard d'années?
Défi temporel: comment illustrer un milliard d'années?  Les trois composants indispensables à l'émergence de la vie
Les trois composants indispensables à l'émergence de la vie  Pourquoi le Genre Homo a frôlé l'extinction il y a 900 000 ans?
Pourquoi le Genre Homo a frôlé l'extinction il y a 900 000 ans?  La première étape vers l'émergence de la vie
La première étape vers l'émergence de la vie  La biosphère de l'ombre
La biosphère de l'ombre  Déclin de l'Anthropocentrisme
Déclin de l'Anthropocentrisme  La limule, un fossile vivant!
La limule, un fossile vivant!  Biosignatures ou présence de vie dans l'Univers
Biosignatures ou présence de vie dans l'Univers  Origine de la vie: Théorie de la panspermie
Origine de la vie: Théorie de la panspermie  Origine de la vie: Théorie des fumeurs blancs
Origine de la vie: Théorie des fumeurs blancs  Pourquoi 37 degrés Celsius?
Pourquoi 37 degrés Celsius?  Sommes-nous seuls dans le cosmos? Entre science et spéculation
Sommes-nous seuls dans le cosmos? Entre science et spéculation  Traces de Vie dans la Glace: L’Émergence des Mammouths Préhistoriques
Traces de Vie dans la Glace: L’Émergence des Mammouths Préhistoriques  Le Dryas: Le mini-âge glaciaire qui faucha la mégafaune
Le Dryas: Le mini-âge glaciaire qui faucha la mégafaune  Les Deux Grandes Glaciations: Survivre dans les Océans d’une Terre Gelée
Les Deux Grandes Glaciations: Survivre dans les Océans d’une Terre Gelée  Régénération chez les Animaux suite à Amputation: La Repousse Organique
Régénération chez les Animaux suite à Amputation: La Repousse Organique  Aux Confins de la Vie: Mephisto, Ver des Profondeurs Infernales
Aux Confins de la Vie: Mephisto, Ver des Profondeurs Infernales  Découverte de buckyballs solides dans l'espace
Découverte de buckyballs solides dans l'espace  La Marche Humaine: Aux Origines de la Bipédie chez les Hominidés
La Marche Humaine: Aux Origines de la Bipédie chez les Hominidés 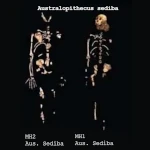 Karabo: Une fenêtre sur l'évolution humaine
Karabo: Une fenêtre sur l'évolution humaine  Entropie: Qu'est-ce que le temps?
Entropie: Qu'est-ce que le temps?  Le passage entre l'inerte et le vivant
Le passage entre l'inerte et le vivant  Le grand récit de la complexité: des particules élémentaires aux premiers organismes
Le grand récit de la complexité: des particules élémentaires aux premiers organismes  Le mégapode utilise la chaleur volcanique
Le mégapode utilise la chaleur volcanique  Ardipithecus: L'hominidé éthiopien de 4,4 millions d’années
Ardipithecus: L'hominidé éthiopien de 4,4 millions d’années  Sélection Naturelle: La Phalène du Bouleau
Sélection Naturelle: La Phalène du Bouleau  L'Ordovicien: L'ère des coraux, des trilobites et des graptolites
L'Ordovicien: L'ère des coraux, des trilobites et des graptolites  L’Eau liquide, Bien Plus Qu’un Solvant: Un Accélérateur de Réactions Chimiques
L’Eau liquide, Bien Plus Qu’un Solvant: Un Accélérateur de Réactions Chimiques  Néandertal: Le Cousin Disparu de l’Humanité
Néandertal: Le Cousin Disparu de l’Humanité  Asimo le futur humanoïde
Asimo le futur humanoïde  Quelles Conditions Ont Permis l’Émergence de la Vie?
Quelles Conditions Ont Permis l’Émergence de la Vie?  Paradoxe de Fermi et caverne de Platon: Sommes-nous seuls ou aveugles?
Paradoxe de Fermi et caverne de Platon: Sommes-nous seuls ou aveugles? 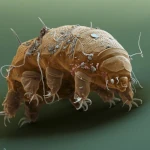 Les tardigrades: Ces créatures indestructibles qui défient les lois de la biologie
Les tardigrades: Ces créatures indestructibles qui défient les lois de la biologie  Toumaï: l'un des plus anciens homininés connus
Toumaï: l'un des plus anciens homininés connus  L'Arbre du Vivant: Des milliards d'espèces disparues et une seule communauté ancestrale
L'Arbre du Vivant: Des milliards d'espèces disparues et une seule communauté ancestrale  La Vie dans les Abysses: L'Extême Adaptation des Créatures
La Vie dans les Abysses: L'Extême Adaptation des Créatures  Cyanobactéries et Crise de l’Oxygène: Une Catastrophe Écologique Primordiale
Cyanobactéries et Crise de l’Oxygène: Une Catastrophe Écologique Primordiale  De la Matière à la Vie: La Frontière Floue de l’Émergence Biologique
De la Matière à la Vie: La Frontière Floue de l’Émergence Biologique  La Plus Petite Grenouille du Monde: Secrets Physiologiques d’un Microvertébré
La Plus Petite Grenouille du Monde: Secrets Physiologiques d’un Microvertébré  L'explication du Petit âge glaciaire
L'explication du Petit âge glaciaire  La Lumière de la Vie: une Biosignature révélée par la Lune
La Lumière de la Vie: une Biosignature révélée par la Lune  Lumière Vivante: Les Secrets Éblouissants de la Bioluminescence
Lumière Vivante: Les Secrets Éblouissants de la Bioluminescence  Au delà de nos sens, les grandes révolutions scientifiques
Au delà de nos sens, les grandes révolutions scientifiques  La Soupe Primitive: Berceau Chimique de la Vie Terrestre
La Soupe Primitive: Berceau Chimique de la Vie Terrestre  Écologie et effondrement: le cas de l’Île de Pâques
Écologie et effondrement: le cas de l’Île de Pâques  Les Fractales: Des structures universelles auto-organisées
Les Fractales: Des structures universelles auto-organisées