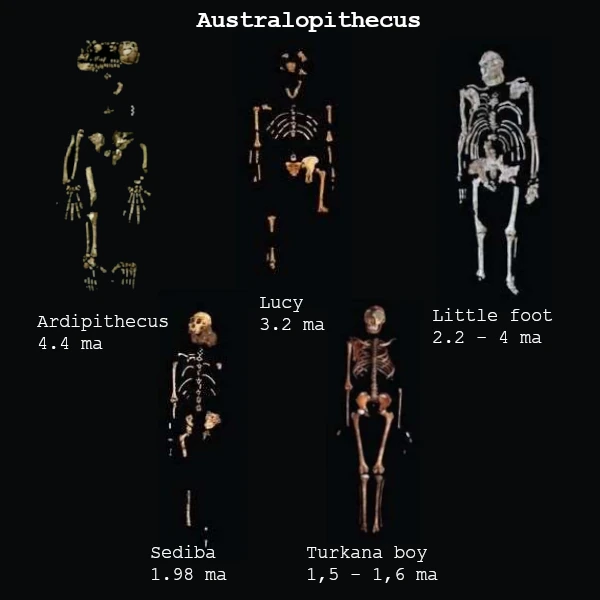
人科包括所有在与黑猩猩谱系分化后与现代人(智人)相关的物种。 古遗传学分析和化石证据表明,这一分化发生在700万至600万年前,正值非洲气候发生重大变革的时期。
最早被确认的代表是乍得沙赫人(约700万年前),发现于乍得。其头骨具有先进的枕骨大孔,表明其具备部分双足行走的姿势。随后是图根原人(约600万年前,肯尼亚)和卡达巴地猿(580万至520万年前),它们已显示出运动适应特征,表明从树栖生活向双足行走的过渡。
距今450万至200万年前,南方古猿成为主要人科动物。著名的"露西"化石所属的阿法南方古猿(390万-300万年前)展现出更显著的双足直立行走特征,但其颅容量(约400立方厘米)仍与大型猿类相近。这些物种与生态多样化共存,栖息地从林地环境到更开阔的热带草原均有分布。
约250万年前,人属的出现标志着进化史上的重大转折点。能人和鲁道夫人与最早的石器(奥杜韦文化)相关联,这体现了新的认知能力。不久后,直立人(约190万年前)成为世界上最早的伟大拓殖者,离开非洲向亚洲和欧洲迁徙,掌握了火的使用,并发展出有组织的社会。
在过去的一百万年里,人类谱系变得更加复杂,出现了海德堡人、尼安德特人、神秘的丹尼索瓦人,以及岛屿物种如弗洛勒斯人和吕宋人。最终,约30万年前在非洲出现的智人,以符号语言、艺术和累积文化的出现为特征,造就了当今全球的多样性与主导地位。
因此,古人类在超过700万年的演化历程中,展现了多样分化与灭绝的复杂过程——多个物种曾共存于世,直至我们这一物种成为人属唯一的幸存者。这种枝状演化表明,人类并非线性进步的产物,而是由自然选择、气候与文化共同塑造的平行谱系交织而成的结果。
| 物种 | 时期(百万年) | 主要地点 | 主要特点 |
|---|---|---|---|
| 乍得沙赫人 | ~7.0 | 乍得 | 扁平颅骨,枕骨大孔位置暗示部分双足行走 |
| 奥罗林·图根猿 | 6.0 | 肯尼亚 | 股骨适应双足行走,原始牙齿 |
| 阿尔迪皮特克斯·卡达巴 | 5.8 – 5.2 | 埃塞俄比亚 | 树栖灵长类与两足动物之间的过渡 |
| 始祖地猿 | 4.4 | 埃塞俄比亚 | 混合树栖与双足行走,具备抓握能力的手 |
| 南方古猿阿法种 | 4.2 – 3.9 | 肯尼亚 | 确认为双足行走,原始齿列 |
| 南方古猿阿法种 | 3.9 – 3.0 | 埃塞俄比亚,坦桑尼亚 | 露西;高效双足行走,颅容量约400立方厘米 |
| 南方古猿普罗米修斯(小脚) | 3.67 | 南非(斯泰克方丹) | 近乎完整的骨架,双足行走与树栖生活 |
| 南方古猿巴赫尔加扎利种 | 3.5 | 乍得 | 发现于远离东非大裂谷的地方,多样的适应性 |
| 南方古猿非洲种 | 3.0 – 2.1 | 南非 | 陶恩儿童头骨,已发展出双足行走能力 |
| 南方古猿加氏种 | 2.5 | 埃塞俄比亚 | 与第一批石器工具相关联 |
| 南方古猿源泉种 | 2.0 | 南非 | 介于南方古猿和人属之间的中间特征 |
| 傍人埃塞俄比亚种 | 2.7 – 2.3 | 肯尼亚 | 黑色颅骨,带有明显的矢状嵴 |
| 傍人鲍氏种 | 2.3 – 1.2 | 东非 | 巨大的颌骨,特化的素食饮食 |
| 傍人罗百氏 | 2.0 – 1.5 | 南非 | 牙齿非常宽大,适应于研磨。 |
| 能人 | 2.4 – 1.6 | 东非 | 奥杜威工具的第一位使用者 |
| 鲁道夫人 | 2.4 – 1.8 | 肯尼亚 | 比能人更大的颅容量 |
| 直立人 | 1.9 – 0.1 | 非洲、亚洲、欧洲 | 走出非洲的第一批伟大迁徙者,掌握了火的使用 |
| 先驱人 | 1.2 – 0.8 | 西班牙 | 尼安德特人与智人可能的共同祖先 |
| 海德堡人 | 0.7 – 0.2 | 欧洲,非洲 | 有组织的猎人,可能是智人和尼安德特人的共同祖先。 |
| 弗洛勒斯人 | 0.1 – 0.06 | 印度尼西亚(弗洛勒斯岛) | “霍比特人”,体型矮小(约1米),脑容量较小 |
| 纳莱迪人 | ~0.3 | 南非 | 可能的仪式性丧葬行为 |
| 吕宋人 | 0.07 | 菲律宾 | 岛屿物种,古老特征的混合体 |
| 尼安德特人 | 0.4 - 0.04 | 欧洲,亚洲 | 复杂的文化,可能发展出了清晰的语言 |
| 龙人 | 0.3 – 0.15 | 中国 | “龙人”,与尼安德特人和丹尼索瓦人相近 |
| 智人 | 0.3 – 至今 | 随后是非洲,再是全球。 | 符号语言、抽象思维、累积文化 |
来源:史密森尼人类起源计划、巴黎国家自然历史博物馆、利基基金会、Berger等人(2015)、Stringer(2022)。
 快速进化:21世纪达尔文式的惊人发现
快速进化:21世纪达尔文式的惊人发现  致命失衡:无限小的潜藏力量
致命失衡:无限小的潜藏力量  演化的伟大过滤器:费米悖论的关键
演化的伟大过滤器:费米悖论的关键  向日葵为何朝向太阳?拉格朗日量给出的答案
向日葵为何朝向太阳?拉格朗日量给出的答案  2026年世界人口:各大洲人口趋势
2026年世界人口:各大洲人口趋势  生命为何源于失衡,又死于热力学平衡
生命为何源于失衡,又死于热力学平衡  电磁波谱与视觉:我们的眼睛如何感知地球
电磁波谱与视觉:我们的眼睛如何感知地球  自我与非我:身份认同的一种简化物理解读
自我与非我:身份认同的一种简化物理解读  分子钟:从随机突变到时间测量
分子钟:从随机突变到时间测量  白沙脚印:美洲最早的人类足迹
白沙脚印:美洲最早的人类足迹 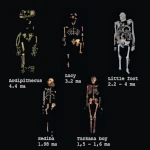 古人类:出现、扩张与灭绝
古人类:出现、扩张与灭绝  重大自然灾害:最可能发生的威胁是什么?
重大自然灾害:最可能发生的威胁是什么?  主要文明崩溃:关键时期与原因
主要文明崩溃:关键时期与原因 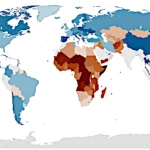 生育率下降:人口灾难还是自然演化?
生育率下降:人口灾难还是自然演化?  自然选择 vs 偶然性:为什么进化不是抽奖?
自然选择 vs 偶然性:为什么进化不是抽奖?  如果生命起源于地球呢?泛种论理论的一场革命
如果生命起源于地球呢?泛种论理论的一场革命  将颠覆世界的重大分岔:生存还是崩溃?
将颠覆世界的重大分岔:生存还是崩溃?  原始化学:最初的有机分子源自何处?
原始化学:最初的有机分子源自何处?  一氧化碳与二氧化碳:两种气体,两种风险,两种生物机制
一氧化碳与二氧化碳:两种气体,两种风险,两种生物机制  自发同步:从物理到生命的普遍现象
自发同步:从物理到生命的普遍现象  时间挑战:如何形象化十亿年?
时间挑战:如何形象化十亿年?  生命涌现的三个基本要素
生命涌现的三个基本要素  为什么90万年前人属差点灭绝?
为什么90万年前人属差点灭绝?  生命涌现的第一步
生命涌现的第一步  影子生物圈
影子生物圈  人类中心主义的衰落
人类中心主义的衰落  鲎,一种活化石!
鲎,一种活化石!  生物印记或宇宙中的生命存在
生物印记或宇宙中的生命存在  地球上生命的起源:泛种论
地球上生命的起源:泛种论  地球上生命的起源:白烟囱理论
地球上生命的起源:白烟囱理论  为什么是37摄氏度?
为什么是37摄氏度?  我们在宇宙中孤独吗?科学与推测之间
我们在宇宙中孤独吗?科学与推测之间  冰层中的生命痕迹:史前猛犸象的出现
冰层中的生命痕迹:史前猛犸象的出现  新仙女木期:消灭巨型动物的小冰期
新仙女木期:消灭巨型动物的小冰期  两大冰期:在冰冻地球的海洋中生存
两大冰期:在冰冻地球的海洋中生存  动物截肢后的再生:组织的再生长
动物截肢后的再生:组织的再生长  生命的极限:墨菲斯托,地狱深渊之虫
生命的极限:墨菲斯托,地狱深渊之虫  太空中发现固体巴基球
太空中发现固体巴基球  人类行走:人科动物双足行走的起源
人类行走:人科动物双足行走的起源 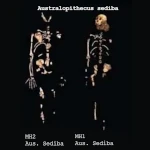 卡拉博:人类演化的一扇窗口
卡拉博:人类演化的一扇窗口  熵:时间是什么?
熵:时间是什么?  从非生命到生命的过渡
从非生命到生命的过渡  复杂性的宏大叙事:从基本粒子到第一批生物
复杂性的宏大叙事:从基本粒子到第一批生物  营冢鸟利用火山热
营冢鸟利用火山热  阿尔迪皮特库斯:440万年前的埃塞俄比亚人科动物
阿尔迪皮特库斯:440万年前的埃塞俄比亚人科动物  自然选择:桦尺蠖
自然选择:桦尺蠖  奥陶纪:珊瑚、三叶虫和笔石的时代
奥陶纪:珊瑚、三叶虫和笔石的时代  液态水,不仅是溶剂,更是化学反应的催化剂
液态水,不仅是溶剂,更是化学反应的催化剂  尼安德特人:人类失落的表亲
尼安德特人:人类失落的表亲  ASIMO,未来的人形机器人
ASIMO,未来的人形机器人  哪些条件促成了生命的出现?
哪些条件促成了生命的出现?  费米悖论与柏拉图洞穴:我们孤独还是盲目?
费米悖论与柏拉图洞穴:我们孤独还是盲目?  水熊虫:挑战生物学法则的坚不可摧的生物
水熊虫:挑战生物学法则的坚不可摧的生物  图迈:已知最古老的人科动物之一
图迈:已知最古老的人科动物之一  生命之树:数十亿灭绝物种与一个共同祖先群体
生命之树:数十亿灭绝物种与一个共同祖先群体  深渊中的生命:生物的极端适应
深渊中的生命:生物的极端适应  蓝藻与氧气危机:一场原始的生态灾难
蓝藻与氧气危机:一场原始的生态灾难  从物质到生命:生物涌现的模糊边界
从物质到生命:生物涌现的模糊边界  世界上最小的青蛙:微型脊椎动物的生理秘密
世界上最小的青蛙:微型脊椎动物的生理秘密  小冰期的解释
小冰期的解释  生命之光:月球揭示的生物印记
生命之光:月球揭示的生物印记  活光:生物发光的炫目秘密
活光:生物发光的炫目秘密  超越感官,伟大的科学革命
超越感官,伟大的科学革命  原始汤:地球生命的化学摇篮
原始汤:地球生命的化学摇篮  世界人口:从十亿人到人口饱和
世界人口:从十亿人到人口饱和  生态与崩溃:复活节岛的案例
生态与崩溃:复活节岛的案例  分形:普遍的自组织结构
分形:普遍的自组织结构