
答案在于劫持一种基本的生物学机制:特定的分子识别(埃米尔·费舍尔的“锁钥模型”)。一个微小的分子,如毒素或毒物,可以完美模仿天然“钥匙”的形状和电子分布。通过以极强的亲和力插入生物“锁”(受体、酶、离子通道),它永久性地阻断或激活一项关键功能,引发连锁反应。因此,肉毒杆菌毒素(1纳克/千克)、氰化物(几毫克/千克)或河豚毒素(2毫克)能麻痹生命功能,这表明原因的规模往往与后果的严重程度毫无关联。
生命有机体是一个复杂且有序的物理化学系统,维持在非平衡稳态中。这种动态组织通过持续的自由能交换得以稳定。其表面上的稳定性依赖于由反馈回路、能量交换和物质流动构成的密集网络。在此背景下,引入一个微小的化学片段可能成为真正的灾难触发因素。当数千个反应相互耦合时,局部扰动可能通过多米诺效应传播、放大,并最终导致生命平衡的全局性崩溃。
注:术语“非平衡”并非指不稳定或混沌状态。它描述的是开放生物系统的稳态——尽管持续产生熵,但宏观属性保持稳定。这种状态通过自由能和物质的持续流动得以维持;若无这些输入,系统将自发向热力学平衡演化,届时所有功能组织将消失殆尽。
生命在其最复杂的形式中,是一种令人惊叹的精密结构。 人体由大约 \( 30 \times 10^{12} \) 个细胞组成,每个细胞包含数十亿个以纳米精度排列的分子。 然而,这台强大的机器却可能因与极微量物质的相互作用而瘫痪、失衡甚至崩溃。 一个“化学微粒”、一个单分子,甚至一个孤立的原子,如何能压倒一个庞大的生物体? 这个问题看似生物学范畴,其终极答案却深藏于原子物理与量子力学的法则中:正是电子云的互补性与分子轨道的能量,决定了某个化学片段是否会与生物靶标结合,从而通过级联反应破坏关键功能。
注:在生物系统中,临界阈值的概念至关重要。低于某一浓度时,分子无害;超过该浓度后,整体动态发生转变,且与增加的量不成正比。
微小力量的力量,源于对基本生物机制的劫持:特异性分子识别,由埃米尔·费歇尔(1852-1919)的“锁钥模型”概念化。这一原理是细胞通信的基石。生物体的蛋白质(受体、酶、离子通道)具有独特电子形状的结合位点,即真正的“分子锁”。只有内源性信号分子——完美适配的“钥匙”——才能与它们结合,触发精确的反应:打开通道、激活酶、调节基因。这是一个超高效的过滤系统,使细胞能够安全、协调地交换信息和流动。
悲剧发生,当一颗“化学微粒”(毒素、毒药或药物)完美模仿了天然钥匙的形状和电子分布。 这把“假钥匙”随后以极强的亲和力插入锁中,将机制永久锁定在“开启”或“关闭”状态。 尽管基于微弱的物理相互作用(氢键、范德华力等),这种被劫持的结合却具有极高的特异性,足以不可逆地瘫痪一项关键的生物功能。 对抗巨人的最强大武器,便是锻造出那把能打开(或永远关闭)其引擎室大门的钥匙。
关于基本机制的说明:现代生物学通过提及立体电子互补性,对费舍尔模型进行了完善。相关的"形状"指的是电子云的形状。这种精确的量子匹配,是数十亿年进化的结果,使得该系统对细胞而言既极其可靠,又悲惨地易受化学劫持。毒性或治疗效果正是源于这种分子身份盗用。
具体实例完美诠释了伊利亚·普里高津(1917-2003)所理论化的耗散结构概念。作为耗散结构的有机体,仅在狭窄的条件范围内保持稳定。以下每个例子都展示了"化学微粒"如何作为微小而精确的"驱动力",将生命功能推离其稳定域,引发功能性的相变——这种相变往往是灾难性的。
| 代理 / 分子 | 尺寸/极小数量 | 生物体内的靶点 | 主要后果与机制 |
|---|---|---|---|
| 肉毒杆菌毒素(A型) | ~1 纳克/千克(估计致死剂量) | 运动神经末梢中的SNARE蛋白 | 弛缓性麻痹。阻断神经递质乙酰胆碱的释放。 |
| 氰化物离子(CN⁻) | 每千克几毫克 | 细胞色素c氧化酶(线粒体呼吸链复合物IV) | 细胞窒息。阻断末端电子传递,停止ATP生成。 |
| 河豚毒素(TTX) | 约2毫克(总致死剂量) | 神经元中的电压门控钠通道 | 神经性麻痹。物理性地阻断通道,阻止动作电位的产生。 |
| 地高辛(洋地黄) | 窄治疗剂量(约0.1毫克/天) | 心肌细胞中的Na⁺/K⁺ ATP酶泵 | 调节心脏力量。抑制泵功能,增加细胞内钙离子浓度和收缩力。 |
| 蓖麻 | 每公斤几微克 | 核糖体RNA的28S亚基(胞质溶胶中) | 蛋白质合成停止。使核糖体RNA残基脱嘌呤,从而使核糖体失活。 |
| 一氧化碳(CO) | 空气中0.1%(长时间暴露) | 血红蛋白中的血红素(氧气结合位点) | 严重缺氧。与血红素结合的亲和力比氧气高200倍,形成碳氧血红蛋白。 |
| 伊马替尼(药物) | 治疗剂量约为400毫克/天 | BCR-ABL酪氨酸激酶(突变型)的ATP位点 | 慢性髓性白血病的缓解。竞争性抑制致癌酶的激活。 |
| 鹅膏菌(死亡帽蘑菇) | ~0.1 mg/kg(致死剂量) | RNA聚合酶II(转录酶) | 肝功能衰竭。阻断基因转录,导致程序性细胞死亡(凋亡)。 |
| 先天性畸形(例如,沙利度胺) | 妊娠期单次剂量 | Cereblon蛋白(调控肢体发育) | 短肢畸形。它使蛋白质复合体发生偏移,导致生长因子的异常降解。 |
| 点突变(例如,镰状细胞贫血) | 在约30亿个核苷酸中替换1个 | HBB基因(编码β-珠蛋白) | 镰状细胞贫血。谷氨酸→缬氨酸的替换,改变了血红蛋白和红细胞的结构。 |
来源:《药理学与实验治疗学杂志》、美国国家医学图书馆(NIH)、欧洲药品管理局(EMA)、《临床毒理学》。
生命是一台具有非凡坚固性和韧性的机器,是数十亿年进化的结果。然而,正是其复杂性使其容易受到微小、有针对性的干扰。理解这一原理,既能揭示剧毒物质的来源,也能阐明精准医学的基础。“生命机器中的微粒”提醒我们,在生命世界中,原因的大小往往与后果的严重程度无关。
由埃米尔·费歇尔(1852-1919)提出的这一模型描述了特定的分子识别机制。人体内的蛋白质(受体、酶)具有独特电子形状的结合位点(锁)。只有内源性信号分子(天然钥匙)才能与之结合,触发精确的生理反应。毒素或药物可通过完美模拟天然钥匙充当"假钥匙",随后将机制永久锁定在"开启"或"关闭"状态。
主要原因是毒性分子对其生物靶点具有高亲和力和高特异性。例如,氰化物(CN⁻)以极高的亲和力结合线粒体呼吸链的复合物IV,从而阻断ATP(细胞能量)的产生。肉毒杆菌毒素(1 ng/kg)作用于神经末梢的SNARE蛋白,阻止神经递质的释放。即使是微乎其微的浓度,也能饱和关键靶点,引发一连串的失效反应。
临界阈值是指分子(有毒物质或药物)达到危险或有效时的浓度。低于此阈值,身体能够解毒、清除或补偿干扰,而不会造成重大后果。高于此阈值,调节机制不堪重负,整体动态会突然发生逆转。这是一种"全有或全无"的概念:微乎其微的额外剂量就能使稳定状态转为灾难,且与添加剂量无直接比例关系。
 快速进化:21世纪达尔文式的惊人发现
快速进化:21世纪达尔文式的惊人发现  致命失衡:无限小的潜藏力量
致命失衡:无限小的潜藏力量  演化的伟大过滤器:费米悖论的关键
演化的伟大过滤器:费米悖论的关键  向日葵为何朝向太阳?拉格朗日量给出的答案
向日葵为何朝向太阳?拉格朗日量给出的答案  2026年世界人口:各大洲人口趋势
2026年世界人口:各大洲人口趋势  生命为何源于失衡,又死于热力学平衡
生命为何源于失衡,又死于热力学平衡  电磁波谱与视觉:我们的眼睛如何感知地球
电磁波谱与视觉:我们的眼睛如何感知地球  自我与非我:身份认同的一种简化物理解读
自我与非我:身份认同的一种简化物理解读  分子钟:从随机突变到时间测量
分子钟:从随机突变到时间测量  白沙脚印:美洲最早的人类足迹
白沙脚印:美洲最早的人类足迹  古人类:出现、扩张与灭绝
古人类:出现、扩张与灭绝  重大自然灾害:最可能发生的威胁是什么?
重大自然灾害:最可能发生的威胁是什么?  主要文明崩溃:关键时期与原因
主要文明崩溃:关键时期与原因  生育率下降:人口灾难还是自然演化?
生育率下降:人口灾难还是自然演化?  自然选择 vs 偶然性:为什么进化不是抽奖?
自然选择 vs 偶然性:为什么进化不是抽奖?  如果生命起源于地球呢?泛种论理论的一场革命
如果生命起源于地球呢?泛种论理论的一场革命  将颠覆世界的重大分岔:生存还是崩溃?
将颠覆世界的重大分岔:生存还是崩溃?  原始化学:最初的有机分子源自何处?
原始化学:最初的有机分子源自何处?  一氧化碳与二氧化碳:两种气体,两种风险,两种生物机制
一氧化碳与二氧化碳:两种气体,两种风险,两种生物机制  自发同步:从物理到生命的普遍现象
自发同步:从物理到生命的普遍现象  时间挑战:如何形象化十亿年?
时间挑战:如何形象化十亿年?  生命涌现的三个基本要素
生命涌现的三个基本要素  为什么90万年前人属差点灭绝?
为什么90万年前人属差点灭绝?  生命涌现的第一步
生命涌现的第一步  影子生物圈
影子生物圈  人类中心主义的衰落
人类中心主义的衰落  鲎,一种活化石!
鲎,一种活化石!  生物印记或宇宙中的生命存在
生物印记或宇宙中的生命存在  地球上生命的起源:泛种论
地球上生命的起源:泛种论  地球上生命的起源:白烟囱理论
地球上生命的起源:白烟囱理论  为什么是37摄氏度?
为什么是37摄氏度?  我们在宇宙中孤独吗?科学与推测之间
我们在宇宙中孤独吗?科学与推测之间  冰层中的生命痕迹:史前猛犸象的出现
冰层中的生命痕迹:史前猛犸象的出现  新仙女木期:消灭巨型动物的小冰期
新仙女木期:消灭巨型动物的小冰期  两大冰期:在冰冻地球的海洋中生存
两大冰期:在冰冻地球的海洋中生存  动物截肢后的再生:组织的再生长
动物截肢后的再生:组织的再生长  生命的极限:墨菲斯托,地狱深渊之虫
生命的极限:墨菲斯托,地狱深渊之虫  太空中发现固体巴基球
太空中发现固体巴基球  人类行走:人科动物双足行走的起源
人类行走:人科动物双足行走的起源  卡拉博:人类演化的一扇窗口
卡拉博:人类演化的一扇窗口  熵:时间是什么?
熵:时间是什么?  从非生命到生命的过渡
从非生命到生命的过渡 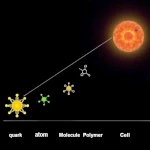 复杂性的宏大叙事:从基本粒子到第一批生物
复杂性的宏大叙事:从基本粒子到第一批生物  营冢鸟利用火山热
营冢鸟利用火山热  阿尔迪皮特库斯:440万年前的埃塞俄比亚人科动物
阿尔迪皮特库斯:440万年前的埃塞俄比亚人科动物  自然选择:桦尺蠖
自然选择:桦尺蠖  奥陶纪:珊瑚、三叶虫和笔石的时代
奥陶纪:珊瑚、三叶虫和笔石的时代  液态水,不仅是溶剂,更是化学反应的催化剂
液态水,不仅是溶剂,更是化学反应的催化剂  尼安德特人:人类失落的表亲
尼安德特人:人类失落的表亲  ASIMO,未来的人形机器人
ASIMO,未来的人形机器人  哪些条件促成了生命的出现?
哪些条件促成了生命的出现?  费米悖论与柏拉图洞穴:我们孤独还是盲目?
费米悖论与柏拉图洞穴:我们孤独还是盲目?  水熊虫:挑战生物学法则的坚不可摧的生物
水熊虫:挑战生物学法则的坚不可摧的生物  图迈:已知最古老的人科动物之一
图迈:已知最古老的人科动物之一  生命之树:数十亿灭绝物种与一个共同祖先群体
生命之树:数十亿灭绝物种与一个共同祖先群体  深渊中的生命:生物的极端适应
深渊中的生命:生物的极端适应  蓝藻与氧气危机:一场原始的生态灾难
蓝藻与氧气危机:一场原始的生态灾难  从物质到生命:生物涌现的模糊边界
从物质到生命:生物涌现的模糊边界  世界上最小的青蛙:微型脊椎动物的生理秘密
世界上最小的青蛙:微型脊椎动物的生理秘密  小冰期的解释
小冰期的解释  生命之光:月球揭示的生物印记
生命之光:月球揭示的生物印记  活光:生物发光的炫目秘密
活光:生物发光的炫目秘密  超越感官,伟大的科学革命
超越感官,伟大的科学革命  原始汤:地球生命的化学摇篮
原始汤:地球生命的化学摇篮  世界人口:从十亿人到人口饱和
世界人口:从十亿人到人口饱和  生态与崩溃:复活节岛的案例
生态与崩溃:复活节岛的案例  分形:普遍的自组织结构
分形:普遍的自组织结构