
その答えは、基本的な生物学的メカニズムである特異的分子認識(エミール・フィッシャーの「鍵と鍵穴」モデル)の乗っ取りにあります。ごく微量な分子、毒素や毒物は、天然の「鍵」の形と電子分布を完全に模倣することができます。驚くべき親和性で生物学的な「鍵穴」(受容体、酵素、イオンチャネル)に挿入されることで、必須の機能を恒久的にブロックまたは活性化し、ドミノ効果を引き起こします。例えば、ボツリヌス毒素(1 ng/kg)、シアン化物(数 mg/kg)、テトロドトキシン(2 mg)は生命維持機能を麻痺させ、原因の規模と結果の大きさがしばしば無関係であることを示しています。
生物は、熱力学的非平衡定常状態に維持された、複雑で組織化された物理化学システムです。 この動的な組織は、自由エネルギーの絶え間ない交換によって安定化されています。 その見かけの安定性は、フィードバック、エネルギー交換、物質の流れの密なネットワークに依存しています。 この文脈において、極小の化学的断片の導入は、文字通りの災害の引き金となる可能性があります。 数千の反応が結合している場合、局所的な攪乱が伝播し、ドミノ効果によって増幅され、生命維持の均衡の全体的な破壊につながる可能性があります。
N.B.:
熱力学的非平衡という表現は、不安定またはカオスな状態を指すものではありません。 これは、開放生物システムに特有の定常状態を記述しており、マクロな性質がエントロピーの継続的な生成にもかかわらず安定したままである状態を指します。 この状態は、自由エネルギーと物質の永続的な流れによって維持されています。これらの供給がなければ、システムは自発的に熱力学的平衡に向かい、すべての機能的組織が消失します。
生命は、その最も進化した形態において、驚異的な複雑さを持つ構造です。 人間は約 \( 30 \times 10^{12} \) 個の細胞から構成され、それぞれの細胞にはナノメートル単位の精度で組織化された数十億の分子が含まれています。 しかし、この驚異的な機械は、極めて微量の物質との相互作用によって麻痺し、不均衡に陥り、打ち倒される可能性があります。 「化学的な砂粒」である1つの分子や単一の原子が、巨大な生物を打ち倒すことができるのはなぜでしょうか? この一見生物学的な問いに対する究極の答えは、原子物理学と量子力学の法則にあります:電子雲の相補性と分子軌道のエネルギーが、化学的断片が生物学的標的に結合し、必須の機能をカスケード的に不均衡にするかどうかを決定するのです。
N.B.:
生物システムにおいて、臨界閾値の概念は中心的です。 ある濃度以下では、分子は無害です。 しかし、それを超えると、全体的なダイナミクスが変化し、追加された量と直接的な比例関係はありません。
極小の力は、基本的な生物学的メカニズムである特異的分子認識の乗っ取りによって説明されます。これはエミール・フィッシャー(1852-1919)の「鍵と錠前」モデルによって概念化されました。 この原理は、細胞間コミュニケーションの基盤です。 生物のタンパク質(受容体、酵素、イオンチャンネル)は、独特の電子形状を持つ結合部位、つまり「分子的な錠前」を持っています。 内因性のシグナル分子、すなわち完全に適合した「鍵」のみが、これらに結合して正確な応答を引き起こすことができます:チャンネルを開く、酵素を活性化する、遺伝子を調節するなど。 これは、細胞が情報や流れを安全かつ協調的に交換できる超効率的なフィルタリングシステムです。
悲劇は、「化学的な砂粒」(毒素、毒、薬)が自然の鍵の形状と電子分布を完全に模倣するときに起こります。 この「偽の鍵」は、恐ろしい親和性で錠前に挿入され、メカニズムを永久的な「オン」または「オフ」状態にロックします。 水素結合やファンデルワールス力などの弱い物理的相互作用に基づいているにもかかわらず、この乗っ取られた結合は非常に特異的であるため、必須の生物学的機能を不可逆的に麻痺させる可能性があります。 したがって、巨人に対する最も強力な武器は、そのエンジンルームのドアを開く(または永遠に閉じる)鍵を鍛えることです。
基本メカニズムに関するN.B.:
現代の生物学は、フィッシャーのモデルを立体電子的相補性という言葉で洗練させています。 関連する「形状」は電子雲のものです。 この正確な量子的マッチングは、数十億年の進化の結果であり、システムを細胞にとって信じられないほど信頼性の高いものにすると同時に、化学的乗っ取りに対して悲劇的に脆弱にします。 毒性または治療効果は、この分子的アイデンティティの簒奪から生じます。
具体的な例は、イリヤ・プリゴジン(1917-2003)が理論化した散逸構造の概念を完璧に示しています。 生物は散逸構造として、狭い条件範囲内でのみ安定です。 以下の各例は、「化学的な砂粒」がどのように無限小ながら正確な「強制」として作用し、生命維持機能をその安定領域から押し出し、しばしば破局的な機能的相転移を引き起こすかを示しています。
| 剤/分子 | サイズ/極微量 | 生物内の標的 | 主要な結果とメカニズム |
|---|---|---|---|
| ボツリヌス毒素(A型) | ~1 kgあたり1ナノグラム(推定致死量) | 運動神経終末のSNAREタンパク質 | 弛緩性麻痺。神経伝達物質アセチルコリンの放出を阻害。 |
| シアン化物イオン (CN⁻) | 1 kgあたり数ミリグラム | ミトコンドリア呼吸鎖複合体IVのシトクロムcオキシダーゼ | 細胞性窒息。電子の最終移動を阻害し、ATP生成を停止。 |
| テトロドトキシン (TTX) (フグ) | ~2 mg(総致死量) | ニューロンの電位依存性ナトリウムチャンネル | 神経麻痺。物理的にチャンネルをブロックし、活動電位の発生を阻害。 |
| ジギトキシン(ジギタリス) | 狭い治療用量(~0.1 mg/日) | 心筋細胞のNa⁺/K⁺ ATPaseポンプ | 心筋力の調節。ポンプを阻害し、細胞内カルシウムと収縮力を増加。 |
| リシン | 1 kgあたり数マイクログラム | リボソームRNAの28Sサブユニット(細胞質内) | タンパク質合成の停止。リボソームRNA残基を脱プリン化し、リボソームを不活化。 |
| 一酸化炭素 (CO) | 空気中の0.1%(長期曝露) | ヘモグロビンのヘム(O₂結合部位) | 重度の低酸素症。酸素の200倍の親和性でヘムに結合し、カルボキシヘモグロビンを形成。 |
| イマチニブ(医薬品) | 治療用量~400 mg/日 | 変異BCR-ABLチロシンキナーゼのATP部位 | 慢性骨髄性白血病の寛解。競合的に腫瘍遺伝子酵素の活性化を阻害。 |
| アマニチン(ドクツルタケ) | ~0.1 mg/kg(致死量) | 転写酵素RNAポリメラーゼII | 肝不全。遺伝子転写を阻害し、プログラム細胞死(アポトーシス)を引き起こす。 |
| 先天性奇形(例:サリドマイド) | 妊娠中の単回投与 | 四肢の発達を調節するセレブロンタンパク質 | アザラシ肢症。タンパク質複合体を逸脱させ、成長因子の異常分解を引き起こす。 |
| 点突然変異 (例:鎌状赤血球症) | ~30億中1ヌクレオチドの置換 | βグロビンをコードするHBB遺伝子 | 鎌状赤血球症。グルタミン酸→バリンの変化により、ヘモグロビンと赤血球の構造が変化。 |
出典:Journal of Pharmacology and Experimental Therapeutics、National Library of Medicine (NIH)、European Medicines Agency (EMA)、臨床毒物学。
生命は、数十億年の進化の結果として、驚異的な堅牢性と回復力を持つ機械です。 しかし、その複雑さゆえに、小さな標的攪乱に対して脆弱です。 この原理を理解することは、恐ろしい毒の源と精密医療の基礎の両方を理解することです。 「生物機械の中の砂粒」は、生命の世界では、原因の規模が結果の大きさとしばしば無関係であることを思い出させてくれます。
エミール・フィッシャー(1852-1919)によって提唱されたこのモデルは、特異的分子認識を説明しています。生体内のタンパク質(受容体、酵素)は、ユニークな電子形状を持つ結合部位(鍵穴)を持っています。正確な応答を引き起こすために結合できるのは、内在性のシグナル分子(天然の鍵)だけです。毒素や薬剤は、天然の鍵を完全に模倣することで「偽の鍵」として作用し、その後、メカニズムを恒久的な「オン」または「オフ」の状態に固定します。
主な理由は、毒分子の生物学的標的に対する高い親和性と特異性です。例えば、シアン化物(CN⁻)はミトコンドリア呼吸鎖の複合体IVに非常に高い親和性で結合し、ATP(細胞エネルギー)の生成を阻害します。ボツリヌス毒素(1 ng/kg)は神経終末のSNAREタンパク質を標的とし、神経伝達物質の放出を防ぎます。ほんの微量な濃度でも必須の標的を飽和させ、障害の連鎖を引き起こす可能性があります。
臨界閾値とは、分子(毒、薬剤)が危険または効果的になる濃度です。この閾値を下回ると、生物は大きな結果もなく、その撹乱を解毒、排除、または補償することができます。それを超えると、調節メカニズムは圧倒され、全体的な動態は突然崩壊します。これは「オール・オア・ナッシング」の概念です:ほんの微量な追加量が、加えられた用量と直接的な比例関係なく、安定した状態を破滅に変える可能性があります。
 致命的不均衡:極小の物質の予想外の力
致命的不均衡:極小の物質の予想外の力  進化の大フィルター:フェルミのパラドックスの鍵
進化の大フィルター:フェルミのパラドックスの鍵  なぜヒマワリは太陽の方を向くのか?ラグランジアンによる解答
なぜヒマワリは太陽の方を向くのか?ラグランジアンによる解答  世界人口2026:大陸別の人口動向
世界人口2026:大陸別の人口動向  なぜ生命は不均衡から生まれ、熱力学的平衡で消滅するのか
なぜ生命は不均衡から生まれ、熱力学的平衡で消滅するのか  電磁スペクトルと視覚:私たちの目が捉える地球の姿
電磁スペクトルと視覚:私たちの目が捉える地球の姿  自己と非自己:物理法則から見たアイデンティティの簡単な解説
自己と非自己:物理法則から見たアイデンティティの簡単な解説  分子時計:突然変異の偶然性から時間の測定へ
分子時計:突然変異の偶然性から時間の測定へ  ホワイトサンズの足跡:アメリカ大陸の最初の一歩
ホワイトサンズの足跡:アメリカ大陸の最初の一歩 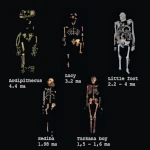 ホミニン:出現、拡散、絶滅
ホミニン:出現、拡散、絶滅 主要な自然災害:最も可能性の高い脅威は何か?
主要な自然災害:最も可能性の高い脅威は何か? 文明の大崩壊:重要な時期と原因
文明の大崩壊:重要な時期と原因 出生率の低下:人口災害か自然な進化か?
出生率の低下:人口災害か自然な進化か? 自然選択 vs 偶然:なぜ進化は宝くじではないのか?
自然選択 vs 偶然:なぜ進化は宝くじではないのか? 生命が地球から始まったら?パンスペルミア理論の革命
生命が地球から始まったら?パンスペルミア理論の革命 世界を激変させる大分岐:生存か崩壊か?
世界を激変させる大分岐:生存か崩壊か? 原始化学:最初の有機分子はどこで生まれたのか?
原始化学:最初の有機分子はどこで生まれたのか? COとCO₂:2つのガス、2つのリスク、2つの生物学的メカニズム
COとCO₂:2つのガス、2つのリスク、2つの生物学的メカニズム 自発的同期:物理学から生命までの普遍的現象
自発的同期:物理学から生命までの普遍的現象 時間的課題:10億年をどのように視覚化するか?
時間的課題:10億年をどのように視覚化するか? 生命の誕生に不可欠な3つの要素
生命の誕生に不可欠な3つの要素 なぜホモ属は90万年前に絶滅の危機に瀕したのか?
なぜホモ属は90万年前に絶滅の危機に瀕したのか? 影の生物圏
影の生物圏 人間中心主義の衰退
人間中心主義の衰退 カブトガニ:生きている化石!
カブトガニ:生きている化石! 宇宙における生命の存在:バイオシグネチャー
宇宙における生命の存在:バイオシグネチャー 人工知能の課題と脅威
人工知能の課題と脅威 生命の起源:パンスペルミア理論
生命の起源:パンスペルミア理論 生命の起源:ホワイトスモーカー理論
生命の起源:ホワイトスモーカー理論 なぜ37度セルシウスなのか?
なぜ37度セルシウスなのか? 私たちは宇宙で孤独なのか?科学と推測の間で
私たちは宇宙で孤独なのか?科学と推測の間で 氷の中の生命の痕跡:先史時代のマムートの出現
氷の中の生命の痕跡:先史時代のマムートの出現 ドリアス期:メガファウナを絶滅させたミニ氷河期
ドリアス期:メガファウナを絶滅させたミニ氷河期 2つの大氷河期:凍った地球の海で生き残る
2つの大氷河期:凍った地球の海で生き残る 動物の切断後の再生:器官の再生
動物の切断後の再生:器官の再生 生命の果て:地底のメフィスト、深淵の虫
生命の果て:地底のメフィスト、深淵の虫 宇宙で固体フラーレンが発見される
宇宙で固体フラーレンが発見される 人間の歩行:原人の二足歩行の起源
人間の歩行:原人の二足歩行の起源 カラボ:人間進化の窓
カラボ:人間進化の窓 エントロピー:時間とは何か?
エントロピー:時間とは何か? 無生物から生命への移行
無生物から生命への移行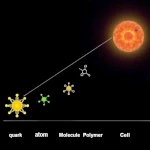 複雑さの物語:素粒子から最初の生物まで
複雑さの物語:素粒子から最初の生物まで メガポード:火山の熱を利用する
メガポード:火山の熱を利用する アルディピテクス:440万年前のエチオピアの原人
アルディピテクス:440万年前のエチオピアの原人 自然選択:カバマダラの例
自然選択:カバマダラの例 オルドビス紀:サンゴ、三葉虫、放散虫の時代
オルドビス紀:サンゴ、三葉虫、放散虫の時代 液体の水:単なる溶媒以上、化学反応の促進剤
液体の水:単なる溶媒以上、化学反応の促進剤 ネアンデルタール人:人類の失われたいとこ
ネアンデルタール人:人類の失われたいとこ アシモ:未来のヒューマノイド
アシモ:未来のヒューマノイド 生命の誕生を可能にした条件は何か?
生命の誕生を可能にした条件は何か? フェルミのパラドックスとプラトンの洞窟:私たちは孤独か、それとも盲目か?
フェルミのパラドックスとプラトンの洞窟:私たちは孤独か、それとも盲目か? クマムシ:生物学の法則に挑戦する不死身の生物
クマムシ:生物学の法則に挑戦する不死身の生物 トゥーマイ:最古の原人の一つ
トゥーマイ:最古の原人の一つ 生命の樹:数十億の絶滅種と単一の祖先コミュニティ
生命の樹:数十億の絶滅種と単一の祖先コミュニティ 深海の生命:極限の適応を遂げた生物
深海の生命:極限の適応を遂げた生物 シアノバクテリアと酸素危機:原始的な環境災害
シアノバクテリアと酸素危機:原始的な環境災害 物質から生命へ:生物学的出現の曖昧な境界
物質から生命へ:生物学的出現の曖昧な境界 世界最小のカエル:微小脊椎動物の生理的秘密
世界最小のカエル:微小脊椎動物の生理的秘密 小氷期の説明
小氷期の説明 生命の光:月が明かすバイオシグネチャー
生命の光:月が明かすバイオシグネチャー 生きている光:生物発光の驚異的な秘密
生きている光:生物発光の驚異的な秘密 感覚を超えて:科学の大革命
感覚を超えて:科学の大革命 原始のスープ:地球生命の化学的揺籃
原始のスープ:地球生命の化学的揺籃 世界人口:10億人から人口飽和へ
世界人口:10億人から人口飽和へ 生態学と崩壊:イースター島の事例
生態学と崩壊:イースター島の事例 フラクタル:自己組織化された普遍的構造
フラクタル:自己組織化された普遍的構造