生命は偶然の産物でも目的の産物でもないとしたら、エネルギーの流れにさらされた物質の相転移の表現なのでしょうか?
無生物の分子から、約38億年前に地球上で生命がどのようにして出現したのでしょうか? この疑問は、最小細胞の原理を調べることで、現代的な視点からアプローチすることができます。 これは、環境の変化に対して調節された内部状態を維持し、自律的に増殖できる最も基本的な組織形態を決定することです。
出典: アレクサンダー・オパーリン(1894年–1980年)やJ.B.S. ホルデイン(1892年–1964年)の古典的な理論では、生命は化学元素の「原始のスープ」から徐々に出現する可能性があると提案されていました。 今日、最小細胞の概念により、生命の誕生を、内部のバランスを維持し自己複製できるシステムが、基本的な物理・化学法則に基づいて自然発生的に形成されるものとして考えることができます。
現代の細胞は、すべての生物(細菌、古細菌、真核生物)の基本単位です。 しかし、最も単純な細胞でさえ、すでに驚異的な複雑さを持つ分子の大聖堂です。
人間の場合、体内には約37兆個の細胞が存在し、200以上の異なる細胞タイプに分かれています。 各細胞タイプは、生物学的機能に適応した特定の形態学的および生化学的特性を持っています。
出典: Geoffrey H. Stoeckiusら(2021年)によると、人間には200以上の異なる細胞タイプが特定されており、それぞれが特定の生理機能に特化しています。 共生細胞説 (リン・マルグリス, 1967年):ミトコンドリアと葉緑体の起源を、真核細胞への細菌の取り込みによって説明しています。
原始地球は、巨大な自然実験の場でした:数兆回の化学反応が同時に数億年にわたって進行しました。 温かい水たまり、熱水噴出孔、多孔質の鉱物や粘土の粒子はそれぞれ、有機分子が集まるミニチュアの実験室でした。 長い時間をかけて自然のサイクルによって絶えず攪拌され、これらの分子は徐々に生命の構成要素(アミノ酸、糖、脂質、RNA塩基)へと至る化学的経路をたどりました。
N.B.:
生命の起源についての議論は活発ですが、一つの合意点があります:アミノ酸はタンパク質を形成し、すべての生物にとって不可欠な分子構成要素です。 さらに、アミノ酸は宇宙空間で自然に形成される可能性があり、隕石からも発見されています。 これは、原始地球上でアミノ酸が利用可能であった可能性を示唆しています。
出典: 原始地球の環境は、熱水噴出孔、温かい水たまり、触媒作用を持つ鉱物が存在し、スタンリー・ミラー(1930年–2007年)とハロルド・ユーリー(1953年)によって「自然の実験室」として前生物的合成の場が提案されました。
原子は、熱運動により絶えず運動しており、分子内でわずかな変位や振動を引き起こします。 水素結合やファンデルワールス力(引力または斥力)などの弱い化学結合が、炭素ベースの原始分子内の原子を結びつけています。 炭素は、4つの安定した共有結合を持つため、分子構造を構築するのに最適な元素です。
分子のランダムな振動は、小さな電気磁石のように振る舞い、化学構造が同時にさまざまな空間的コンフォメーションを探索できるようにします。 新しい分子形態が絶えず生成され、安定した配置の可能性が高まります。 この多様性は、事実上無限の分子形態の貯蔵庫であり、電磁的に安定な構造のみが自然選択にさらされるのに十分長く残るでしょう。
出典: マックス・デルブリュック(1949年)の生物システムにおける確率的変動に関する研究が示すように、これらのランダムな運動により、化学構造が同時に多くのコンフォメーションを試すことが可能になります。 分子自己組織化:ジャン=マリー・レーン(1939年生まれ)による超分子化学の研究は、弱い相互作用(水素結合、ファンデルワールス力)に焦点を当てています。
特定の分子構成は、自己複製を可能にします。これは、構造的相補性という単純な原理に基づいています。
高分子(3D形状に折りたたまれた単鎖)は、型として機能し、その形状と電荷が周囲の自由な化学元素を正確に引き付けます。 これらの自由な元素は、化学的な互換性に従って鎖に沿って配列し、第二の相補的な鎖(写真のネガのようなもの)を形成します。 これにより、周囲の成分からピースごとに組み立てられた、二本の鎖が対になった構造が得られます。
水素結合によって一緒に保たれている二本の鎖は、最終的に環境の熱運動によって自然に分離します。 このエネルギーバランスは重要です。出会いと分離を可能にするのに十分な運動が必要ですが、組み立てられた構造(温かい水たまり、熱水噴出孔など)の完全性を保つために、あまりにも多くの運動は必要ありません。 この相補的な高分子は、元の配列を再現するための型として機能することができます。
この純粋に物理化学的なメカニズム(引き付け→配列→対合→分離→元の配列の復元)によって、原始地球上で自己複製可能なRNA高分子が自然発生的に出現した可能性が理解できます。
出典: RNA分子の自然発生的な複製は、ウォルター・ギルバート(1986年)によって提案されたRNAワールド仮説の枠組みに位置付けられます。この仮説は、RNAが情報を蓄積すると同時に化学反応を触媒することができたことを示唆しています。
システムは自己触媒的になります:一部のRNA分子は他よりも速く複製され、自由なヌクレオチド(A、G、C、U)をより効率的に取り込みます。 小さな誤りや変異が新しい形態を生み出し、複製速度や安定性が異なります。 最も速く安定した分子が繁栄し、効率の悪いものは消えます。 この選択は自然なものであり、物質の絶え間ない流れによって攪拌され供給される非平衡システムにおける化学反応速度論の法則から直接生じます。
出典: 自己触媒的選択のダイナミクスは、アイゲンとシュスター(1977年)によって提案されたハイパーサイクルのモデルを思い起こさせます。このモデルは、競争と複製が非平衡化学システムにおいて特定の分子を優位にする過程を説明しています。
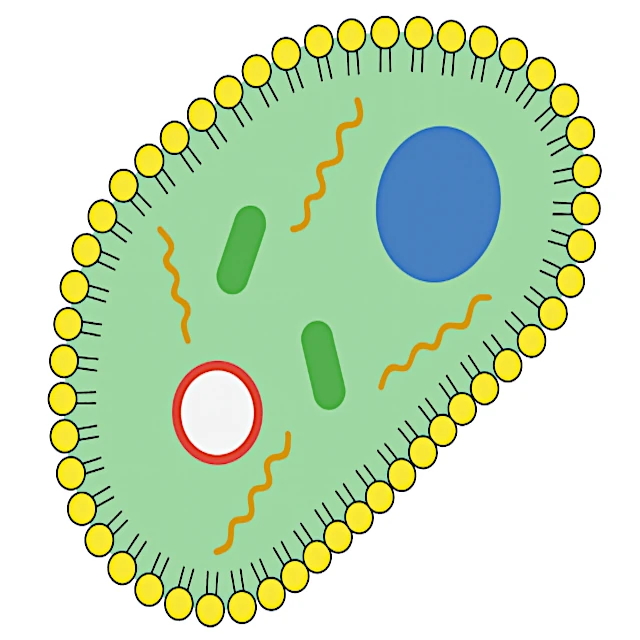
複製可能な分子から真の細胞への移行には、膜区画の出現が必要です。 生物学的膜は、リン脂質、つまり両親媒性分子で構成されており、水中で自然に二重層を形成します。 この自己組織化は自然なものであり、外部からの介入なしに熱力学的に有利に進行します。石鹸の泡が水面で自然に形成されるのと似ています。
脂質膜に保護され、自己複製可能なRNA分子を含む原始細胞は、最小限の生物の基本的な特性を備えています。 代謝(内部成分の生産)、増殖能力(細胞分裂)、伝達可能な情報担体(RNA)を持っています。 この段階から、ダーウィン進化が完全に作用し、環境内で成長し分裂するのに最も効率的な原始細胞を優先します。
出典: リン脂質の二重層への自己組織化は実験的に観察され、両親媒性システムの熱力学によって説明されています (バンガムら, 1965年)。 これは、膜の形成が生物学的な介入を必要としないことを示しています。
 最小作用の原理:なぜ自然は常に最も経済的な道を選ぶのか?
最小作用の原理:なぜ自然は常に最も経済的な道を選ぶのか?  収斂進化:偶然、必然…それとも幻?
収斂進化:偶然、必然…それとも幻?  五大大量絶滅:これらの地球規模の災厄は何を語るのか?
五大大量絶滅:これらの地球規模の災厄は何を語るのか?  氷圏:氷の巨人がまだ私たちを守っている
氷圏:氷の巨人がまだ私たちを守っている  窒素循環:空気から生命へ
窒素循環:空気から生命へ  ジェット気流:大気の川が氾濫するとき
ジェット気流:大気の川が氾濫するとき  炭素サイクル:海洋、森林、地下の調和的な相互作用
炭素サイクル:海洋、森林、地下の調和的な相互作用  水のパラドックス:水なしに生命なし
水のパラドックス:水なしに生命なし  なぜ220V交流がコンセントに?物理学の問題
なぜ220V交流がコンセントに?物理学の問題  最後の氷河期ショック:ヤンガードライアスと先史時代の社会への影響
最後の氷河期ショック:ヤンガードライアスと先史時代の社会への影響  536年:夏のない年と後期古代小氷期の始まり
536年:夏のない年と後期古代小氷期の始まり 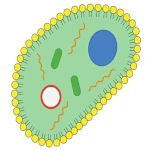 最小細胞:避けられない出現
最小細胞:避けられない出現 

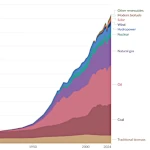 世界のエネルギー消費の進化:蒸気機関からデジタルサーバーまで
世界のエネルギー消費の進化:蒸気機関からデジタルサーバーまで  人口転換:成長か衰退か?
人口転換:成長か衰退か? 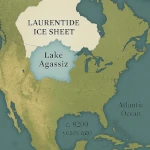 アガシーズ湖:気候を変えた大洪水
アガシーズ湖:気候を変えた大洪水  酔っ払いの森:永久凍土の融解現象を理解する
酔っ払いの森:永久凍土の融解現象を理解する  空気中の電気!
空気中の電気! 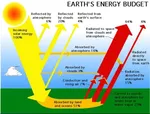 なぜCO2は地面に落ちないのか?
なぜCO2は地面に落ちないのか?  エルニーニョとラニーニャ:太平洋の二つの息吹が地球の気候を律する
エルニーニョとラニーニャ:太平洋の二つの息吹が地球の気候を律する  絶滅危惧種:絶滅のダイナミクス
絶滅危惧種:絶滅のダイナミクス  酸性雨の有毒な遺産
酸性雨の有毒な遺産  気候変動の数字:科学データが語るもの
気候変動の数字:科学データが語るもの  持続可能な開発とは何か?
持続可能な開発とは何か?  アラル海:人類への教訓
アラル海:人類への教訓  三峡ダム:生態系への影響
三峡ダム:生態系への影響 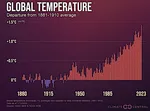 観測史上最も暑い年
観測史上最も暑い年  今後の気候変動の規模
今後の気候変動の規模  危機に瀕する地球:崩壊か再生か?
危機に瀕する地球:崩壊か再生か?  危機に瀕する氷:北極海氷の避けられない減少
危機に瀕する氷:北極海氷の避けられない減少  地球の水資源:海洋から地下水まで
地球の水資源:海洋から地下水まで  海面上昇の速度はどれくらいか?
海面上昇の速度はどれくらいか?  もう一つの気候の脅威:地球が影と湿気に包まれる
もう一つの気候の脅威:地球が影と湿気に包まれる  サハラの年齢:緑の草原から石の砂漠へ
サハラの年齢:緑の草原から石の砂漠へ  1800年から2100年までの世界人口
1800年から2100年までの世界人口  石油:再生可能エネルギー時代における巨人の没落
石油:再生可能エネルギー時代における巨人の没落  カムチャツカのタラバガニ:海の巨人
カムチャツカのタラバガニ:海の巨人  イースター島:社会の崩壊
イースター島:社会の崩壊  世界のエコロジカル・フットプリントのわずかな進展
世界のエコロジカル・フットプリントのわずかな進展 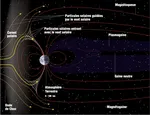 南大西洋異常
南大西洋異常  地盤沈下現象
地盤沈下現象  なぜ地球はこんなに頻繁に曇っているのか?
なぜ地球はこんなに頻繁に曇っているのか?  地球の新しい雲
地球の新しい雲 デザーテック:ヨーロッパとアフリカのための太陽エネルギーの夢
デザーテック:ヨーロッパとアフリカのための太陽エネルギーの夢