チャールズ・ダーウィン(1809-1882)以来、進化論は自然選択の原則に基づいています。 これは、環境に最も適応した形質の伝達を促進します。 一般的なイメージでは、進化は複雑性の増大への線形的な進歩として認識されています。 しかし、自然は進化に特権的な方向性がないことを示しています:進化は単純化された形態、器官の喪失、さらには真の退化に至ることもあります。
進化には方向性も目的もありません。進化は環境が提供する可能性を探求し、機能するものを保持します。たとえそれが見かけ上の単純化であってもです。スティーブン・ジェイ・グールド(1941-2002)が指摘したように、複雑性は生命の偶発的な結果であり、目的地ではありません。 退化は、自然が「進歩」するのではなく、適応することを思い出させます。
N.B.:
退化という用語は、現代生物学によって正式な概念として認められていません。これは、機能的な喪失や進化的な単純化の過程を比喩的に説明するものです。
彼の理論「自然選択による種の起源、あるいは生存競争における優れた種の保存」(1859年)において、チャールズ・ダーウィンは「進化」という言葉を一度も使用しませんでした。 この用語は、最後の版(1872年の第6版)の最終文でのみ登場します: 「生命のこの見方には壮大さがある… これほど単純な始まりから、無数の最も美しく驚異的な形態が進化し、今も進化し続けている。」
ダーウィンは「進化」という言葉を避けていました。なぜなら、彼以前は、この言葉が主に予めプログラムされた発展、内部的な計画の展開(特にラマルクの場合)を指していたからです。しかし、彼の理論は計画や方向性の欠如に基づいていたのです。
知性、サイズ、完全性への継続的な上昇は、人間中心主義に染まったビジョンであり、チャールズ・ダーウィンの理論の誤解です。 時には、最良の適応は退行することです。
退化は「退歩」という否定的な意味での言葉ではありません。 むしろ、生物が複雑な特徴を失い、より単純な形態を獲得する進化的な現象を指します。これは祖先への「戻り」ではなく、減算による新たな適応です。駆動力は「退歩」ではなく、複雑性が負担となるときに単純性を選択する選択圧です。
すべての複雑な構造にはコストがかかります。 各器官、細胞ネットワーク、または発現した遺伝子は、エネルギーを消費し、遺伝的制御とメンテナンスを必要とします。 環境が特定の機能を要求しなくなると、それらを維持していた選択圧が消失します。 種は、生物学的構造を単純化することでエネルギー効率を高めます。
| 生物 | 失われたまたは単純化された特徴 | 適応の原因または文脈 | コメント |
|---|---|---|---|
| 洞窟魚 Astyanax mexicanus | 目の喪失と色素の喪失 | 完全な暗闇での生活、エネルギーの節約 | 視力を失うことは進化の「誤り」ではありません。これは、これらの種が極端な生態的ニッチを征服することを可能にした注目すべき適応です。 |
| 条虫 (Taenia solium) | 消化管の消失 | 宿主からの栄養素の直接吸収 | 代謝の極端な低下と消化器系の喪失は、寄生生活への完全な特化につながり、寄生虫は宿主から直接栄養素を吸収することで生存します。 |
| ヘビ (トカゲの子孫) | 前肢 後肢 | 掘削への適応 体の波動による移動 | 肢の喪失は、掘削生活や体の波動による移動と関連しており、これは穴を掘ったり獲物を追いかけたりするのにより効率的です。 |
| クジラ (Balaenoptera musculus) | 後肢の喪失 | 水生環境への完全な適応 | 後肢は抵抗の原因となり、泳ぎをはるかに非効率にします。 |
| ペンギン (Aptenodytes forsteri) | 飛行能力の喪失 | 翼のヒレへの変化 | 空力から水力への変換により、水中での効率的な推進が可能になります。 |
| ダチョウ (Struthio camelus) | 飛行不能 | 陸上での高速走行への適応 | エネルギーは走行に振り向けられます:残存する翼はバランスと求愛ディスプレイに使用されます。 |
| ハキリアリ Atta cephalotes | セルロースを 消化する能力 | 菌類との共生 分業 | セルロースの消化を共生菌に外部化したため、この生理的能力を失いました。これにより、各コロニーのメンバーが相互扶助的な食物システムに貢献する集団的な特化が促進されます。 |
| 鳥 Apteryx australis (キウイ) | 翼と目の縮小 | ニュージーランドの森林での夜行性および陸上生活 | 羽毛は毛皮のようなふわふわした質感に退化し、一方、末端の鼻孔と過剰に発達した嗅覚がこの単純化を補い、夜行性の捕食者として特化しています。 |
| 両生類 Proteus anguinus | 機能的な目の喪失 | 石灰岩洞窟での地下生活 | 萎縮した視覚器官は光に対する皮膚の感受性に置き換えられました。 |
自然は、意図や方向性なしに、原始的な細胞から複雑な多細胞生物へと、生きている物質の組織化をどのように進めるのでしょうか?
その答えは、開放系の熱力学と自己組織化の論理にあります。
生物学的進化は進歩ではなく、可能性の探求です。 進化には目的がありません(複雑性へ向かう目標はありません)。 各生物学的変化は、単に局所的な制約の結果です:ランダムな突然変異、物理的および化学的な相互作用、そして与えられた環境における自然選択。 これらの制約の一部は、安定した構造の出現、つまりより組織化された構造を促進します。
したがって、私たちが生物圏で観察する複雑性の増大は、普遍的な傾向ではなく、散逸系の物理学の副次的な効果です。
生きているシステムは、熱力学的平衡から遠く離れた開放系です。 それは環境と物質やエネルギーを絶えず交換しています。 イリヤ・プリゴジン(1917-2003)の理論によれば、これらのシステムは、エネルギーの流れが一定の臨界しきい値を超えると、自己組織化することができます。
N.B.:
原理:一定のエネルギーの流れは、エントロピーが外部に散逸する限り、秩序立った構造を維持することができます。
真核細胞への移行は「進歩」ではなく、安定した共生の結果です。 原始的な細胞(古細菌)が、好気性細菌を取り込み、それがミトコンドリアになりました。 この内部共生の過程により、エネルギーをより効率的に利用できるようになり、自己組織化の能力が向上しました。 これは階層的な移行よりもエネルギー的な移行です。
類似した細胞が協力してエネルギーと栄養素の流れをよりよく管理すると、機能的な分化が自然に出現します。 一部の細胞は構造に特化し、他の細胞は繁殖やコミュニケーションに特化します。
組織の各レベル(細胞→組織→器官→生物)は、「計画」の産物ではなく、相互作用の徐々な安定化の結果です。 システムがエネルギーを交換し、情報(遺伝的、エピジェネティック、または化学的)を維持するほど、システムは構造化することができ、動的なバランスを失うことなくなります。 物理的な観点からは、秩序立った複雑な構造を維持するには、一定のエネルギーの流れが必要です。
\( \text{複雑性} \approx \text{安定性} + \text{エネルギーの流れ} + \text{保存された情報} \)
 最小作用の原理:なぜ自然は常に最も経済的な道を選ぶのか?
最小作用の原理:なぜ自然は常に最も経済的な道を選ぶのか?  収斂進化:偶然、必然…それとも幻?
収斂進化:偶然、必然…それとも幻?  五大大量絶滅:これらの地球規模の災厄は何を語るのか?
五大大量絶滅:これらの地球規模の災厄は何を語るのか?  氷圏:氷の巨人がまだ私たちを守っている
氷圏:氷の巨人がまだ私たちを守っている  窒素循環:空気から生命へ
窒素循環:空気から生命へ  ジェット気流:大気の川が氾濫するとき
ジェット気流:大気の川が氾濫するとき  炭素サイクル:海洋、森林、地下の調和的な相互作用
炭素サイクル:海洋、森林、地下の調和的な相互作用 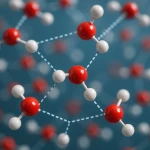 水のパラドックス:水なしに生命なし
水のパラドックス:水なしに生命なし  なぜ220V交流がコンセントに?物理学の問題
なぜ220V交流がコンセントに?物理学の問題  最後の氷河期ショック:ヤンガードライアスと先史時代の社会への影響
最後の氷河期ショック:ヤンガードライアスと先史時代の社会への影響  536年:夏のない年と後期古代小氷期の始まり
536年:夏のない年と後期古代小氷期の始まり  最小細胞:避けられない出現
最小細胞:避けられない出現 

 世界のエネルギー消費の進化:蒸気機関からデジタルサーバーまで
世界のエネルギー消費の進化:蒸気機関からデジタルサーバーまで  人口転換:成長か衰退か?
人口転換:成長か衰退か?  アガシーズ湖:気候を変えた大洪水
アガシーズ湖:気候を変えた大洪水  酔っ払いの森:永久凍土の融解現象を理解する
酔っ払いの森:永久凍土の融解現象を理解する  空気中の電気!
空気中の電気!  なぜCO2は地面に落ちないのか?
なぜCO2は地面に落ちないのか?  エルニーニョとラニーニャ:太平洋の二つの息吹が地球の気候を律する
エルニーニョとラニーニャ:太平洋の二つの息吹が地球の気候を律する  絶滅危惧種:絶滅のダイナミクス
絶滅危惧種:絶滅のダイナミクス  酸性雨の有毒な遺産
酸性雨の有毒な遺産  気候変動の数字:科学データが語るもの
気候変動の数字:科学データが語るもの  持続可能な開発とは何か?
持続可能な開発とは何か?  アラル海:人類への教訓
アラル海:人類への教訓  三峡ダム:生態系への影響
三峡ダム:生態系への影響  観測史上最も暑い年
観測史上最も暑い年  今後の気候変動の規模
今後の気候変動の規模  危機に瀕する地球:崩壊か再生か?
危機に瀕する地球:崩壊か再生か?  危機に瀕する氷:北極海氷の避けられない減少
危機に瀕する氷:北極海氷の避けられない減少  地球の水資源:海洋から地下水まで
地球の水資源:海洋から地下水まで  海面上昇の速度はどれくらいか?
海面上昇の速度はどれくらいか?  もう一つの気候の脅威:地球が影と湿気に包まれる
もう一つの気候の脅威:地球が影と湿気に包まれる  サハラの年齢:緑の草原から石の砂漠へ
サハラの年齢:緑の草原から石の砂漠へ  1800年から2100年までの世界人口
1800年から2100年までの世界人口  石油:再生可能エネルギー時代における巨人の没落
石油:再生可能エネルギー時代における巨人の没落  カムチャツカのタラバガニ:海の巨人
カムチャツカのタラバガニ:海の巨人  イースター島:社会の崩壊
イースター島:社会の崩壊  世界のエコロジカル・フットプリントのわずかな進展
世界のエコロジカル・フットプリントのわずかな進展  南大西洋異常
南大西洋異常  地盤沈下現象
地盤沈下現象  なぜ地球はこんなに頻繁に曇っているのか?
なぜ地球はこんなに頻繁に曇っているのか?  地球の新しい雲
地球の新しい雲 デザーテック:ヨーロッパとアフリカのための太陽エネルギーの夢
デザーテック:ヨーロッパとアフリカのための太陽エネルギーの夢