
物理的および化学的に、水は次のように分類されます。ミネラル、つまり、分子 \(\mathrm{H_2O}\) で構成される単純な無機物質です。有機栄養素(炭水化物、脂質、タンパク質)とは異なり、水は化学エネルギーや細胞が直接使用できる構造要素を提供しません。
したがって、その主な機能は細胞に供給することではなく、細胞を構成することです。細胞内および細胞外環境生活に欠かせないもの。これは万能溶媒としての役割を果たし、栄養素の輸送、老廃物の除去、体温調節、生化学反応の仲介を促進します。
一方、水は、生理学的機能に関与する溶解イオン(ミネラル、電解質)を提供しますが、これらは水自体からではなく、他のミネラルの溶解から得られます。要約すると、水は生命にとって不可欠な媒介物ですが、直接的な栄養供給にはなりません。
生物系では、水は酵素触媒作用において基本的な役割を果たします。酵素の活性ポケットは部分的に水和されていることがよくあります。水は閉じ込められ、配向され、機能化されます。それは電荷の移動を可能にし、励起状態を安定させ、時には試薬または補因子として機能します。
光合成における水の光分解反応や細胞呼吸(電子鎖)において、水は単なる飾りではなく、エネルギー代謝の主役です。
水は単なる万能溶媒ではありません。それは、生命システムと水性環境の化学において積極的な役割を果たします。その極性のおかげで、その水素結合のネットワークは永久に再配置されます (10111 秒あたりの再組織化数)、および遷移状態を安定化するその能力により、液体の水は実際の物質として機能します。化学反応促進剤。
多くの有機溶媒 (ヘキサンやトルエンなど) とは異なり、水は反応物質と直接相互作用し、化学反応の過程を変化させます。
水は主に次の 3 つの方法で反応に影響を与えます。
水は単なる「観客」ではなく、さまざまなレベルで化学反応を形成する積極的な参加者です。
要約すると、不活性溶媒とは異なり、水は次のことができます。反応機構に直接関与する: プロトンの移動 (酸塩基反応)、イオン中間体の安定化、または遷移状態のエネルギー状況の変更によって。それは反応速度論だけでなく、反応の選択性や熱力学にも影響を与えます。
微視的なスケールでは、液体の水は水素結合の三次元ネットワークを形成し、常に再組織化します。結晶秩序とガス状の無秩序の間のこの中間構造は、反応微小環境これにより、反応物間の効率的な衝突が促進されます。
例 1: 水の自動イオン化
水の自動イオン化は、一時的な水素橋を介した 2 つの分子間のプロトンの移動に基づいています。
$$2 \, \mathrm{H_2O} \ \rightleftharpoons \ \mathrm{H_3O^+} + \mathrm{OH^-}$$
この移動はグロッタス機構によって促進されます。グロッタス機構では、水素結合の急速な再構成のおかげで、陽子はそれ自体を移動することなく格子を通って伝播します。この現象は、水などの高度に構造化された移動性の液体媒体中にのみ存在します。
例 2: 水性媒体中でのディールス・アルダー反応
ジエンとジエノフィルとの間のディールス・アルダー反応は、有機溶媒と比較して水中では著しく促進されます。この速度の向上は、疎水効果: 非極性試薬は水によってグループ化され、そのネットワークから疎水性化合物を排除する傾向があります。この凝集圧力により、効果的な衝突の確率が高まり、反応が特定の生成物に向けられ、化学反応性に対する水の構造効果が実証されています。
| 反応 | 溶媒 | 速度定数(k) | コメント |
|---|---|---|---|
| HClのイオン化 | 水の中 | 非常に高い | イオンの極性と水和によって促進される |
| エステル加水分解 | 水の中 | 103倍速 | 求核剤としての水の積極的な関与 |
| 置換SN1 | 水 vs ヘキサン | 水中で加速 | 炭素カチオン中間体の安定化 |
| プロトン移動 | 水 | ほぼ瞬時に | 自己タンパク質分解効果 H2O⇌H3○++OH− |
出典: J. Phys. のデータ。化学。 B (ACS)、アンジュー。ケミーインターナショナル編、F. フランクス著、*Water: A Matrix of Life*。
水は、反応媒体および生命の維持として欠かせない一連のユニークな物理化学的特性を持っています。その高い極性、大きな双極子モーメント (≈1.85 D)、高密度でダイナミックな水素結合ネットワークを形成する優れた能力、広範囲の地上温度に適合する融点と沸点、および高い誘電率 (≈78.5 ~ 25 °C) です。 イオン解離を促進します。
これらの特性の組み合わせにより、流動性と構造の間、効率的な溶媒和と分子運動性の間のまれなバランスが確保され、水が万能溶媒と活性触媒の両方として機能することが可能になります。さらに、分子量が低く粘度が低いため、迅速な輸送と効率的な分子相互作用が促進されます。
液体アンモニア (NH₃)、メタノール (CH₃OH)、ギ酸 (HCOOH) などの他の分子も、極限状態での代替溶媒として考慮されてきましたが、水の重要な特性をすべて兼ね備えたものはありません。たとえば、液体アンモニアは極性が低く、誘電率が低く、水素ネットワークがあまり発達していません。さらに、これらの溶媒は、寿命に適合する温度または圧力範囲がはるかに制限されています。
結論として、他の液体は理論的には水の役割の一部を果たすことができますが、水を複雑な化学や生物学にとって例外的な媒体にする物理化学的特性をすべて備えている既知の分子はありません。これが、今日に至るまで、水が陸上環境においてかけがえのない分子であり続けている理由です。
 致命的不均衡:極小の物質の予想外の力
致命的不均衡:極小の物質の予想外の力  進化の大フィルター:フェルミのパラドックスの鍵
進化の大フィルター:フェルミのパラドックスの鍵  なぜヒマワリは太陽の方を向くのか?ラグランジアンによる解答
なぜヒマワリは太陽の方を向くのか?ラグランジアンによる解答  世界人口2026:大陸別の人口動向
世界人口2026:大陸別の人口動向  なぜ生命は不均衡から生まれ、熱力学的平衡で消滅するのか
なぜ生命は不均衡から生まれ、熱力学的平衡で消滅するのか  電磁スペクトルと視覚:私たちの目が捉える地球の姿
電磁スペクトルと視覚:私たちの目が捉える地球の姿  自己と非自己:物理法則から見たアイデンティティの簡単な解説
自己と非自己:物理法則から見たアイデンティティの簡単な解説  分子時計:突然変異の偶然性から時間の測定へ
分子時計:突然変異の偶然性から時間の測定へ  ホワイトサンズの足跡:アメリカ大陸の最初の一歩
ホワイトサンズの足跡:アメリカ大陸の最初の一歩  ホミニン:出現、拡散、絶滅
ホミニン:出現、拡散、絶滅 主要な自然災害:最も可能性の高い脅威は何か?
主要な自然災害:最も可能性の高い脅威は何か? 文明の大崩壊:重要な時期と原因
文明の大崩壊:重要な時期と原因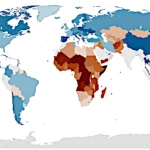 出生率の低下:人口災害か自然な進化か?
出生率の低下:人口災害か自然な進化か? 自然選択 vs 偶然:なぜ進化は宝くじではないのか?
自然選択 vs 偶然:なぜ進化は宝くじではないのか? 生命が地球から始まったら?パンスペルミア理論の革命
生命が地球から始まったら?パンスペルミア理論の革命 世界を激変させる大分岐:生存か崩壊か?
世界を激変させる大分岐:生存か崩壊か? 原始化学:最初の有機分子はどこで生まれたのか?
原始化学:最初の有機分子はどこで生まれたのか? COとCO₂:2つのガス、2つのリスク、2つの生物学的メカニズム
COとCO₂:2つのガス、2つのリスク、2つの生物学的メカニズム 自発的同期:物理学から生命までの普遍的現象
自発的同期:物理学から生命までの普遍的現象 時間的課題:10億年をどのように視覚化するか?
時間的課題:10億年をどのように視覚化するか? 生命の誕生に不可欠な3つの要素
生命の誕生に不可欠な3つの要素 なぜホモ属は90万年前に絶滅の危機に瀕したのか?
なぜホモ属は90万年前に絶滅の危機に瀕したのか? 影の生物圏
影の生物圏 人間中心主義の衰退
人間中心主義の衰退 カブトガニ:生きている化石!
カブトガニ:生きている化石! 宇宙における生命の存在:バイオシグネチャー
宇宙における生命の存在:バイオシグネチャー 人工知能の課題と脅威
人工知能の課題と脅威 生命の起源:パンスペルミア理論
生命の起源:パンスペルミア理論 生命の起源:ホワイトスモーカー理論
生命の起源:ホワイトスモーカー理論 なぜ37度セルシウスなのか?
なぜ37度セルシウスなのか? 私たちは宇宙で孤独なのか?科学と推測の間で
私たちは宇宙で孤独なのか?科学と推測の間で 氷の中の生命の痕跡:先史時代のマムートの出現
氷の中の生命の痕跡:先史時代のマムートの出現 ドリアス期:メガファウナを絶滅させたミニ氷河期
ドリアス期:メガファウナを絶滅させたミニ氷河期 2つの大氷河期:凍った地球の海で生き残る
2つの大氷河期:凍った地球の海で生き残る 動物の切断後の再生:器官の再生
動物の切断後の再生:器官の再生 生命の果て:地底のメフィスト、深淵の虫
生命の果て:地底のメフィスト、深淵の虫 宇宙で固体フラーレンが発見される
宇宙で固体フラーレンが発見される 人間の歩行:原人の二足歩行の起源
人間の歩行:原人の二足歩行の起源 カラボ:人間進化の窓
カラボ:人間進化の窓 エントロピー:時間とは何か?
エントロピー:時間とは何か? 無生物から生命への移行
無生物から生命への移行 複雑さの物語:素粒子から最初の生物まで
複雑さの物語:素粒子から最初の生物まで メガポード:火山の熱を利用する
メガポード:火山の熱を利用する アルディピテクス:440万年前のエチオピアの原人
アルディピテクス:440万年前のエチオピアの原人 自然選択:カバマダラの例
自然選択:カバマダラの例 オルドビス紀:サンゴ、三葉虫、放散虫の時代
オルドビス紀:サンゴ、三葉虫、放散虫の時代 液体の水:単なる溶媒以上、化学反応の促進剤
液体の水:単なる溶媒以上、化学反応の促進剤 ネアンデルタール人:人類の失われたいとこ
ネアンデルタール人:人類の失われたいとこ アシモ:未来のヒューマノイド
アシモ:未来のヒューマノイド 生命の誕生を可能にした条件は何か?
生命の誕生を可能にした条件は何か? フェルミのパラドックスとプラトンの洞窟:私たちは孤独か、それとも盲目か?
フェルミのパラドックスとプラトンの洞窟:私たちは孤独か、それとも盲目か? クマムシ:生物学の法則に挑戦する不死身の生物
クマムシ:生物学の法則に挑戦する不死身の生物 トゥーマイ:最古の原人の一つ
トゥーマイ:最古の原人の一つ 生命の樹:数十億の絶滅種と単一の祖先コミュニティ
生命の樹:数十億の絶滅種と単一の祖先コミュニティ 深海の生命:極限の適応を遂げた生物
深海の生命:極限の適応を遂げた生物 シアノバクテリアと酸素危機:原始的な環境災害
シアノバクテリアと酸素危機:原始的な環境災害 物質から生命へ:生物学的出現の曖昧な境界
物質から生命へ:生物学的出現の曖昧な境界 世界最小のカエル:微小脊椎動物の生理的秘密
世界最小のカエル:微小脊椎動物の生理的秘密 小氷期の説明
小氷期の説明 生命の光:月が明かすバイオシグネチャー
生命の光:月が明かすバイオシグネチャー 生きている光:生物発光の驚異的な秘密
生きている光:生物発光の驚異的な秘密 感覚を超えて:科学の大革命
感覚を超えて:科学の大革命 原始のスープ:地球生命の化学的揺籃
原始のスープ:地球生命の化学的揺籃 世界人口:10億人から人口飽和へ
世界人口:10億人から人口飽和へ 生態学と崩壊:イースター島の事例
生態学と崩壊:イースター島の事例 フラクタル:自己組織化された普遍的構造
フラクタル:自己組織化された普遍的構造