
エチオピアのアファール地方で発見されたアルディピテクス・ラミドゥスは、20 世紀の最も重要な古人類学的発見の 1 つです。最初の化石の破片は 1992 年にティム・ホワイト率いるチームによって発掘されましたが、「アルディ」と呼ばれる部分骨格が特定されたのは 1994 年になってからでした。 440万年前に遡るこの原人は、最初の二足歩行の原人が従来考えられていたようなサバンナではなく、森林環境に生息していました。
アルディピテクスは、原始的な特徴と派生した特徴の魅力的な組み合わせを示します。 化石には、身長約1.20メートル、体重50キロの生物が示されている。 その脳の大きさはおよそ \(300 \text{ to } 350 \text{ cm}^3\) で、現代のチンパンジーの脳に匹敵しました。その歯は大型類人猿の歯とは異なり、犬歯が減少しており、攻撃的ではない社会的行動を示唆しています。広い骨盤と大後頭孔の位置は二足歩行への適応を示しているが、その足には木登り用の対向母趾 (つかみやすい母趾) が残されていた。
二足歩行の出現が開けたサバンナ環境であるとするこれまでの理論とは異なり、アルディピテクスは森林環境で進化した。この発見は、二足歩行を促進する進化の圧力の再考につながりました。二足歩行は、点在する木々の間の移動を容易にしたり、混合環境で食物を輸送したりするために出現した可能性があります。したがって、アルディピテクスは、樹上移動と地上移動を組み合わせたライフスタイルへの顕著な適応を表しました。
アルディピテクス・ラミドゥス、古い440万年、は古代のヒト科であるだけでなく、亜科にも属します。ヒト族つまり、オランウータンよりも人間に近い系統です。
この分類は、アルディピテクスがチンパンジーや人間と共通の祖先を共有しているが、私たちの系統に直接つながる枝上にあることを意味します。時折現れる二足歩行と樹上での能力を組み合わせたそのユニークなキャラクターの組み合わせは、ヒト族の特徴、特に二足歩行の漸進的な進化を示しています。
アルディピテクスは人類として、私たちの系統とチンパンジーの系統の間の分岐の初期段階に関する重要な手がかりを提供し、古代の大型類人猿とより最近のアウストラロピテクスとの間の移行に光を当てます。部族における彼の立場ヒト科それは人類の起源を理解する上で不可欠な要点となります。
注::
あヒト科人間を含む現生および化石のすべての大型類人猿を一堂に集めます。人類オランウータンを除き、ゴリラ、チンパンジー、人間を含む亜科を指します。したがって、すべてのヒト族はヒト族ですが、すべてのヒト族がヒト族であるわけではありません。
アルディピテクス・ラミドゥスは、アウストラロピテクス・アファレンシス(有名な「ルーシー") 約 100 万年。これは、\(5 \text{ から } 7 \text{ 百万年}\) の間と推定される、ヒトとチンパンジーの系統の分岐に近い位置にあります。この年代順の位置により、後の人類につながった適応の重要な証拠となっています。最近の発見は、アルディピテクスがおそらくホモ サピエンスの直接の祖先ではなく、むしろ従兄弟の枝であることを示唆しています。 別の進化の道を模索しました。
| 種 | 年齢 (数百万年) | 頭蓋容量 | 中型 | 平均体重 | 移動モード | ダイエット | 環境 |
|---|---|---|---|---|---|---|---|
| アルディピテクス・ラミドゥス | 4.4 | 300~350cm3 | ≈ 1.20メートル | ≈ 50kg | 時々二足歩行 + 樹上生活 | 雑食性(果物、種子、根、小型無脊椎動物) | 開けた森林と樹林帯 |
| アウストラロピテクス・アナメンシス | 4.2~3.9 | ≈ 365 ~ 370 cm3 | ≈ 1.20 ~ 1.40 m | ≈ 45 ~ 55 kg | 二足歩行は、アルディピテクス、それでも登れます。 | 塊茎と種子を持つ優勢な果食動物 | 開けた森林環境とサバンナエリア |
| アウストラロピテクス・アファレンシス(ルーシー) | 3.2 | 400 ~ 450 cm3 | ≈ 1.05 ~ 1.50 m | ≈ 30 ~ 45 kg | 主張された二足歩行、残りの登り | 主に果食性で、塊茎によって補足される | 東アフリカの樹木サバンナ |
| ホモ・ハビリス | 2.4~1.6 | 600 ~ 700 cm3 | ≈ 1.25 ~ 1.40 m | ≈ 35 ~ 50 kg | 完全二足歩行、石器 | 肉の摂取量が増える雑食動物 | 東アフリカのサバンナと広々とした風景 |
| ホモ・エレクトス | 1.9~0.1 | 850 ~ 1100 cm3 | ≈ 1.50 ~ 1.80 m | ≈ 50 ~ 70 kg | 現代の二足歩行 | 日和見的な雑食性、制御された射撃 | アフリカ、その後ユーラシア (さまざまな気候) |
| ホモ・サピエンス | 0.3 – 現在 | 1300 ~ 1600 cm3 | ≈ 1.60 ~ 1.85 m | ≈ 55 ~ 90 kg | 現代の二足歩行 | 雑食動物、農業、繁殖、象徴文化 | すべての地理的エリア、高い適応能力 |
出典: White T. et al. (2009)、科学。スミソニアン博物館の人類の起源。タタソール I. (2012)、マスターズ・オブ・ザ・プラネット、パルグレイブ・マクミラン。アントン S.C. (2003)、ホモ・エレクトスの自然史。
 進化の大フィルター:フェルミのパラドックスの鍵
進化の大フィルター:フェルミのパラドックスの鍵  なぜヒマワリは太陽の方を向くのか?ラグランジアンによる解答
なぜヒマワリは太陽の方を向くのか?ラグランジアンによる解答  世界人口2026:大陸別の人口動向
世界人口2026:大陸別の人口動向  なぜ生命は不均衡から生まれ、熱力学的平衡で消滅するのか
なぜ生命は不均衡から生まれ、熱力学的平衡で消滅するのか  電磁スペクトルと視覚:私たちの目が捉える地球の姿
電磁スペクトルと視覚:私たちの目が捉える地球の姿  自己と非自己:物理法則から見たアイデンティティの簡単な解説
自己と非自己:物理法則から見たアイデンティティの簡単な解説  分子時計:突然変異の偶然性から時間の測定へ
分子時計:突然変異の偶然性から時間の測定へ  ホワイトサンズの足跡:アメリカ大陸の最初の一歩
ホワイトサンズの足跡:アメリカ大陸の最初の一歩 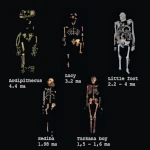 ホミニン:出現、拡散、絶滅
ホミニン:出現、拡散、絶滅 主要な自然災害:最も可能性の高い脅威は何か?
主要な自然災害:最も可能性の高い脅威は何か? 文明の大崩壊:重要な時期と原因
文明の大崩壊:重要な時期と原因 生成AI vs AGI:模倣の終わり、意識の始まりはどこか?
生成AI vs AGI:模倣の終わり、意識の始まりはどこか? 出生率の低下:人口災害か自然な進化か?
出生率の低下:人口災害か自然な進化か? 自然選択 vs 偶然:なぜ進化は宝くじではないのか?
自然選択 vs 偶然:なぜ進化は宝くじではないのか? 生命が地球から始まったら?パンスペルミア理論の革命
生命が地球から始まったら?パンスペルミア理論の革命 世界を激変させる大分岐:生存か崩壊か?
世界を激変させる大分岐:生存か崩壊か? 原始化学:最初の有機分子はどこで生まれたのか?
原始化学:最初の有機分子はどこで生まれたのか? COとCO₂:2つのガス、2つのリスク、2つの生物学的メカニズム
COとCO₂:2つのガス、2つのリスク、2つの生物学的メカニズム 自発的同期:物理学から生命までの普遍的現象
自発的同期:物理学から生命までの普遍的現象 人工ネットワーク vs 生物学的ネットワーク:2つのシステム、共通のアーキテクチャ
人工ネットワーク vs 生物学的ネットワーク:2つのシステム、共通のアーキテクチャ 人間の脳と人工知能:類似点と相違点
人間の脳と人工知能:類似点と相違点 時間的課題:10億年をどのように視覚化するか?
時間的課題:10億年をどのように視覚化するか? 生命の誕生に不可欠な3つの要素
生命の誕生に不可欠な3つの要素 なぜホモ属は90万年前に絶滅の危機に瀕したのか?
なぜホモ属は90万年前に絶滅の危機に瀕したのか? AlphaGo vs AlphaGo Zero:人工知能の革命
AlphaGo vs AlphaGo Zero:人工知能の革命 知的機械の次のステップ
知的機械の次のステップ 生命誕生への第一歩
生命誕生への第一歩 生物学的ニューロンから形式ニューロンへ:脳の単純化
生物学的ニューロンから形式ニューロンへ:脳の単純化 影の生物圏
影の生物圏 人間中心主義の衰退
人間中心主義の衰退 人工知能:巨大化の爆発
人工知能:巨大化の爆発 人工知能が狂ったとき!
人工知能が狂ったとき! 人工知能の誕生:知能の幻想か、本当の知能か?
人工知能の誕生:知能の幻想か、本当の知能か? カブトガニ:生きている化石!
カブトガニ:生きている化石! 宇宙における生命の存在:バイオシグネチャー
宇宙における生命の存在:バイオシグネチャー 人工知能の課題と脅威
人工知能の課題と脅威 機械は人間と同様に言語を理解し、解釈し、生成する方法
機械は人間と同様に言語を理解し、解釈し、生成する方法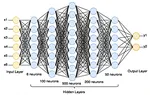 人工ニューラルネットワークの仕組み
人工ニューラルネットワークの仕組み 生命の起源:パンスペルミア理論
生命の起源:パンスペルミア理論 生命の起源:ホワイトスモーカー理論
生命の起源:ホワイトスモーカー理論 なぜ37度セルシウスなのか?
なぜ37度セルシウスなのか? 私たちは宇宙で孤独なのか?科学と推測の間で
私たちは宇宙で孤独なのか?科学と推測の間で 氷の中の生命の痕跡:先史時代のマムートの出現
氷の中の生命の痕跡:先史時代のマムートの出現 ドリアス期:メガファウナを絶滅させたミニ氷河期
ドリアス期:メガファウナを絶滅させたミニ氷河期 2つの大氷河期:凍った地球の海で生き残る
2つの大氷河期:凍った地球の海で生き残る 動物の切断後の再生:器官の再生
動物の切断後の再生:器官の再生 生命の果て:地底のメフィスト、深淵の虫
生命の果て:地底のメフィスト、深淵の虫 宇宙で固体フラーレンが発見される
宇宙で固体フラーレンが発見される 人間の歩行:原人の二足歩行の起源
人間の歩行:原人の二足歩行の起源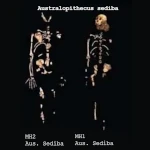 カラボ:人間進化の窓
カラボ:人間進化の窓 過ぎ去る時間
過ぎ去る時間 無生物から生命への移行
無生物から生命への移行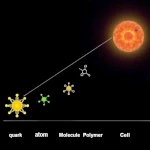 複雑さの物語:素粒子から最初の生物まで
複雑さの物語:素粒子から最初の生物まで メガポード:火山の熱を利用する
メガポード:火山の熱を利用する アルディピテクス:440万年前のエチオピアの原人
アルディピテクス:440万年前のエチオピアの原人 自然選択:カバマダラの例
自然選択:カバマダラの例 オルドビス紀:サンゴ、三葉虫、放散虫の時代
オルドビス紀:サンゴ、三葉虫、放散虫の時代 液体の水:単なる溶媒以上、化学反応の促進剤
液体の水:単なる溶媒以上、化学反応の促進剤 ネアンデルタール人:人類の失われたいとこ
ネアンデルタール人:人類の失われたいとこ アシモ:未来のヒューマノイド
アシモ:未来のヒューマノイド 生命の誕生を可能にした条件は何か?
生命の誕生を可能にした条件は何か? フェルミのパラドックスとプラトンの洞窟:私たちは孤独か、それとも盲目か?
フェルミのパラドックスとプラトンの洞窟:私たちは孤独か、それとも盲目か? クマムシ:生物学の法則に挑戦する不死身の生物
クマムシ:生物学の法則に挑戦する不死身の生物 トゥーマイ:最古の原人の一つ
トゥーマイ:最古の原人の一つ 生命の樹:数十億の絶滅種と単一の祖先コミュニティ
生命の樹:数十億の絶滅種と単一の祖先コミュニティ 深海の生命:極限の適応を遂げた生物
深海の生命:極限の適応を遂げた生物 シアノバクテリアと酸素危機:原始的な環境災害
シアノバクテリアと酸素危機:原始的な環境災害 物質から生命へ:生物学的出現の曖昧な境界
物質から生命へ:生物学的出現の曖昧な境界 世界最小のカエル:微小脊椎動物の生理的秘密
世界最小のカエル:微小脊椎動物の生理的秘密 小氷期の説明
小氷期の説明 生命の光:月が明かすバイオシグネチャー
生命の光:月が明かすバイオシグネチャー 生きている光:生物発光の驚異的な秘密
生きている光:生物発光の驚異的な秘密 感覚を超えて:科学の大革命
感覚を超えて:科学の大革命 原始のスープ:地球生命の化学的揺籃
原始のスープ:地球生命の化学的揺籃 世界人口:10億人から人口飽和へ
世界人口:10億人から人口飽和へ 生態学と崩壊:イースター島の事例
生態学と崩壊:イースター島の事例 フラクタル:自己組織化された普遍的構造
フラクタル:自己組織化された普遍的構造