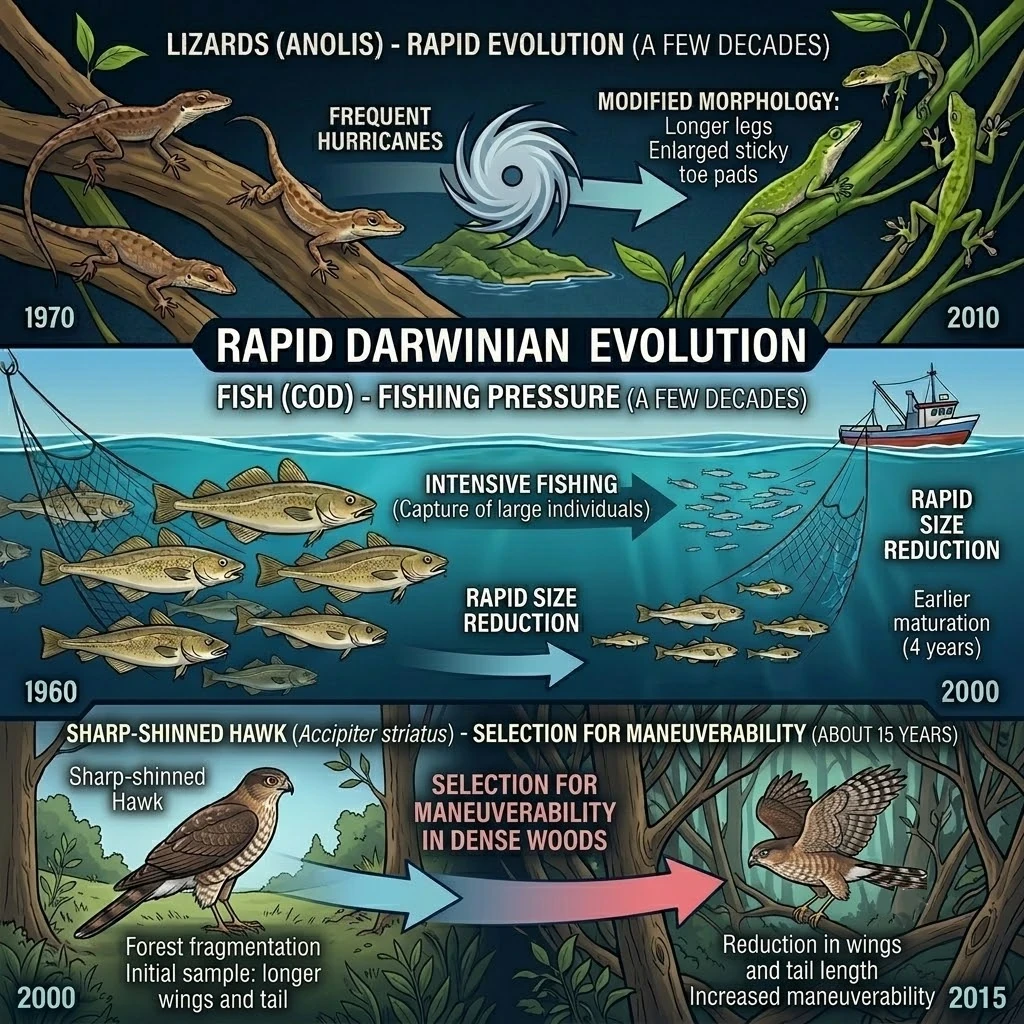
チャールズ・ダーウィン(1809-1882)が考えていたように、自然選択は数百万年にわたる緩慢なプロセスであるという通念に反して、進化は実際には非常に速いものです。 なぜでしょうか?3つの条件が進化の速度を加速させるからです:強い選択圧(環境の急激な変化)、個体群内の高い遺伝的多様性、そして短い世代時間です。 これらの要因が揃うと、わずか数世代で大きな変化が目に見える形で現れます。 例えば、蚊が数シーズンで殺虫剤に耐性を持つようになったり、魚が20年程度で漁業の圧力により体が小さくなったり、壁トカゲが30年で消化器系を変化させたりするのです。 これらの発見はダーウィンの理論に反するのではなく、むしろそれを豊かにします:進化は一様に遅いわけではなく、状況が要求する時には加速することもあるのです。
種の起源(1859年)において、ダーウィンは変化の遅さを強調しています:「自然選択は、軽微で連続的な変異を蓄積することによってのみ作用する」。 彼にとって、化石記録に移行形態が存在しないことは、それらの記録の不完全さによって説明できましたが、彼は一様で非常に遅いリズムを固く信じていました。 この視点は系統漸進主義と呼ばれ、1世紀以上にわたって進化生物学を支配してきました。
しかしながら、1970年代にはすでに、ナイルズ・エルドレッジ(1943–2022)とスティーブン・ジェイ・グールド(1941–2002)が断続平衡理論を提唱しました:進化は長い停滞期と急速な種分化のエピソードを繰り返すというものです。 今日、分子生物学と個体群生態学は、選択圧が強い場合(気候変動、人間の影響、汚染、捕食)には進化は非常に速く進行することを確認しています。 これらの発見はダーウィンの理論に反するのではなく、進化のリズムが極めて変動的であることを示すことで、その理論を豊かにしています。
世代時間が数分または数時間で測られる微生物において、進化が速いことは容易に理解できます。 しかし、より驚くべきことは、この進化の加速現象が大型生物(魚、爬虫類、鳥、哺乳類)でも観察されることです。以下の例がそれを証明しています。 個体群が高速で進化するためには、4つの要素が必要です:
このように、加速した進化は自然選択の壮大な実例です:それは無から何かを創造するのではなく、既に存在するものを選別し増幅させるのです。時には驚くべき速さで。 ダーウィンが予想しなかったのは、人間が環境をどれほど速く変化させるか、そしてそれが極めて強い選択圧を生み出すかということです。
強い環境圧と自然選択は、形態的、生理的、または行動的な変化を地質学的瞬間に引き起こす可能性があります。 環境圧は直接突然変異を加速させるわけではありません。 しかしながら、それは既に個体群内に存在する有利な突然変異の選択と固定を促進します。 例えば、人間では、各世代で約70の新しい突然変異が発生します。これは安定した数字ですが、種全体のスケールでは、各世代で何十億もの新しいバリエーションを生み出すことになります。
| 種 | 環境 / 圧力 | 観察された変化 | 時間スケール |
|---|---|---|---|
| イタリアトカゲ (Podarcis sicula) | 1971年にクロアチアのポド・ムルチャル島に導入、植物性の食事が増加 | 盲腸(結腸弁)の発達、植物を消化するため、頭部が広くなり、噛む力が強くなる | 30年未満(約30世代) |
| ハチクマ (Accipiter fuscus) | 森林の断片化、機動性に関する選択 | 密集した木々の間を飛ぶため、翼と尾の長さが短縮 | 約15年 |
| レマン湖のパーチ | 大型個体を選択的に捕獲する集中的な漁業 | 捕獲される前に繁殖するため、性成熟が早まり、体サイズが小さくなる | 20年(1980年代から2000年代) |
| ダーウィンフィンチ (Geospiza fortis) | ガラパゴス諸島の極度の干ばつ(1977年) | 硬い種子を食べるため、くちばしのサイズが5%増加 | 2年(1回の干ばつ) |
| ハマナガイナガ (Biston betularia) | イギリスの産業汚染、木の幹の黒ずみ | 汚れた幹によりよく擬態するため、明るい型から暗い型(メラニック)への置き換え | 50年(19世紀半ばから20世紀初頭) |
| 大西洋サケ (Salmo salar) | 大型で成熟した個体を選択的に捕獲する集中的な漁業 | 成魚のサイズが小さくなり、性成熟が早まる、文書化された遺伝的変化 | 30年(1970年代から2000年) |
| アフリカゾウ (Loxodonta africana) | 大型の牙を持つ個体を選択的に狩猟する密猟 | 牙のない個体の割合が大幅に増加(X染色体に関連する遺伝的形質) | 30年(モザンビーク内戦、1977-1992年) |
| 汚染された川の魚(メダカ、ウグイ) | カドミウム、銅、亜鉛などの重金属を含む産業廃棄物 | メタロチオネインをコードする遺伝子の発現変化を伴う重金属に対する遺伝的耐性の発達 | 数十年 |
ダーウィンの進化論は、変動的なリズムを持ち、選択の強さと人口統計学的特性によって決まります。 ダーウィンが人間のスケールではほとんど認識できないプロセスと考えていたものが、特定の条件下ではわずか数年で劇的に変化する可能性があります。 この再発見(加速する進化)はダーウィンの理論に反するのではなく、それを現代化し豊かにします。 そして何よりも、生命は静的なものではないことを思い出させてくれます:生命は、我々が与える撹乱に反応し、時には驚異的な速さで適応するのです。 これは魅力的でありながらも不安を感じさせる事実であり、地球のための我々の決断を導くべきです。
スティーブン・ジェイ・グールド(1941-2002)が言ったように:「進化には目的はないが、結果はある。」
ダーウィンは、自然選択が微小な変異に作用し、地質学的な時間スケールを必要とすると思っていました。彼はメカニズムについては正しかったのですが、短い世代と極端な圧力に関連する急速な進化の可能性については知りませんでした。今日、我々は進化が非常に速い(数世代)ことができることを知っていますが、これはダーウィンの漸進主義に反するものではありません:進化は既存の遺伝的多様性を利用するのです。ダーウィンは単に変化する環境における可能性のある速度を過小評価していたのです。
現代の進化とは、人間の時間スケールで測定可能な進化的変化(数十年、またはそれ以下)を指します。これはしばしば急速な環境変化(汚染、気候、捕食、漁業)によって引き起こされます。例としては、抗生物質耐性、鳥のくちばしのサイズの変化、植物の重金属耐性などがあります。
いいえ。急速な進化には、短い世代時間、高い遺伝的多様性、強い選択圧が必要です。大型哺乳類、数百年を生きる木、または個体数の少ない種は、はるかにゆっくりと適応します。そのため、現在の絶滅は主にこれらのゆっくりと更新される種に影響を与えています。
短い生命周期と高い遺伝的多様性を持つ種(昆虫、一年草、小型魚)については、急速な適応が可能です。しかし、長寿の象徴的な種(ホッキョクグマ、サンゴ、ウミガメ)については、進化が現在の気候変動のペースについていくにはあまりに遅すぎます。したがって、優先事項は依然として排出量の削減と生息地の保護です。
 21世紀のダーウィンの雷鳴:加速する進化
21世紀のダーウィンの雷鳴:加速する進化  致命的不均衡:極小の物質の予想外の力
致命的不均衡:極小の物質の予想外の力  進化の大フィルター:フェルミのパラドックスの鍵
進化の大フィルター:フェルミのパラドックスの鍵  なぜヒマワリは太陽の方を向くのか?ラグランジアンによる解答
なぜヒマワリは太陽の方を向くのか?ラグランジアンによる解答  世界人口2026:大陸別の人口動向
世界人口2026:大陸別の人口動向  なぜ生命は不均衡から生まれ、熱力学的平衡で消滅するのか
なぜ生命は不均衡から生まれ、熱力学的平衡で消滅するのか  電磁スペクトルと視覚:私たちの目が捉える地球の姿
電磁スペクトルと視覚:私たちの目が捉える地球の姿  自己と非自己:物理法則から見たアイデンティティの簡単な解説
自己と非自己:物理法則から見たアイデンティティの簡単な解説  分子時計:突然変異の偶然性から時間の測定へ
分子時計:突然変異の偶然性から時間の測定へ  ホワイトサンズの足跡:アメリカ大陸の最初の一歩
ホワイトサンズの足跡:アメリカ大陸の最初の一歩 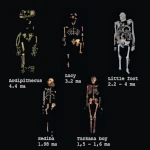 ホミニン:出現、拡散、絶滅
ホミニン:出現、拡散、絶滅 主要な自然災害:最も可能性の高い脅威は何か?
主要な自然災害:最も可能性の高い脅威は何か? 文明の大崩壊:重要な時期と原因
文明の大崩壊:重要な時期と原因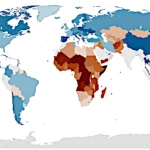 出生率の低下:人口災害か自然な進化か?
出生率の低下:人口災害か自然な進化か? 自然選択 vs 偶然:なぜ進化は宝くじではないのか?
自然選択 vs 偶然:なぜ進化は宝くじではないのか? 生命が地球から始まったら?パンスペルミア理論の革命
生命が地球から始まったら?パンスペルミア理論の革命 世界を激変させる大分岐:生存か崩壊か?
世界を激変させる大分岐:生存か崩壊か? 原始化学:最初の有機分子はどこで生まれたのか?
原始化学:最初の有機分子はどこで生まれたのか? COとCO₂:2つのガス、2つのリスク、2つの生物学的メカニズム
COとCO₂:2つのガス、2つのリスク、2つの生物学的メカニズム 自発的同期:物理学から生命までの普遍的現象
自発的同期:物理学から生命までの普遍的現象 時間的課題:10億年をどのように視覚化するか?
時間的課題:10億年をどのように視覚化するか? 生命の誕生に不可欠な3つの要素
生命の誕生に不可欠な3つの要素 なぜホモ属は90万年前に絶滅の危機に瀕したのか?
なぜホモ属は90万年前に絶滅の危機に瀕したのか? 影の生物圏
影の生物圏 人間中心主義の衰退
人間中心主義の衰退 カブトガニ:生きている化石!
カブトガニ:生きている化石! 宇宙における生命の存在:バイオシグネチャー
宇宙における生命の存在:バイオシグネチャー 人工知能の課題と脅威
人工知能の課題と脅威 生命の起源:パンスペルミア理論
生命の起源:パンスペルミア理論 生命の起源:ホワイトスモーカー理論
生命の起源:ホワイトスモーカー理論 なぜ37度セルシウスなのか?
なぜ37度セルシウスなのか? 私たちは宇宙で孤独なのか?科学と推測の間で
私たちは宇宙で孤独なのか?科学と推測の間で 氷の中の生命の痕跡:先史時代のマムートの出現
氷の中の生命の痕跡:先史時代のマムートの出現 ドリアス期:メガファウナを絶滅させたミニ氷河期
ドリアス期:メガファウナを絶滅させたミニ氷河期 2つの大氷河期:凍った地球の海で生き残る
2つの大氷河期:凍った地球の海で生き残る 動物の切断後の再生:器官の再生
動物の切断後の再生:器官の再生 生命の果て:地底のメフィスト、深淵の虫
生命の果て:地底のメフィスト、深淵の虫 宇宙で固体フラーレンが発見される
宇宙で固体フラーレンが発見される 人間の歩行:原人の二足歩行の起源
人間の歩行:原人の二足歩行の起源 カラボ:人間進化の窓
カラボ:人間進化の窓 エントロピー:時間とは何か?
エントロピー:時間とは何か? 無生物から生命への移行
無生物から生命への移行 複雑さの物語:素粒子から最初の生物まで
複雑さの物語:素粒子から最初の生物まで メガポード:火山の熱を利用する
メガポード:火山の熱を利用する アルディピテクス:440万年前のエチオピアの原人
アルディピテクス:440万年前のエチオピアの原人 自然選択:カバマダラの例
自然選択:カバマダラの例 オルドビス紀:サンゴ、三葉虫、放散虫の時代
オルドビス紀:サンゴ、三葉虫、放散虫の時代 液体の水:単なる溶媒以上、化学反応の促進剤
液体の水:単なる溶媒以上、化学反応の促進剤 ネアンデルタール人:人類の失われたいとこ
ネアンデルタール人:人類の失われたいとこ アシモ:未来のヒューマノイド
アシモ:未来のヒューマノイド 生命の誕生を可能にした条件は何か?
生命の誕生を可能にした条件は何か? フェルミのパラドックスとプラトンの洞窟:私たちは孤独か、それとも盲目か?
フェルミのパラドックスとプラトンの洞窟:私たちは孤独か、それとも盲目か? クマムシ:生物学の法則に挑戦する不死身の生物
クマムシ:生物学の法則に挑戦する不死身の生物 トゥーマイ:最古の原人の一つ
トゥーマイ:最古の原人の一つ 生命の樹:数十億の絶滅種と単一の祖先コミュニティ
生命の樹:数十億の絶滅種と単一の祖先コミュニティ 深海の生命:極限の適応を遂げた生物
深海の生命:極限の適応を遂げた生物 シアノバクテリアと酸素危機:原始的な環境災害
シアノバクテリアと酸素危機:原始的な環境災害 物質から生命へ:生物学的出現の曖昧な境界
物質から生命へ:生物学的出現の曖昧な境界 世界最小のカエル:微小脊椎動物の生理的秘密
世界最小のカエル:微小脊椎動物の生理的秘密 小氷期の説明
小氷期の説明 生命の光:月が明かすバイオシグネチャー
生命の光:月が明かすバイオシグネチャー 生きている光:生物発光の驚異的な秘密
生きている光:生物発光の驚異的な秘密 感覚を超えて:科学の大革命
感覚を超えて:科学の大革命 原始のスープ:地球生命の化学的揺籃
原始のスープ:地球生命の化学的揺籃 世界人口:10億人から人口飽和へ
世界人口:10億人から人口飽和へ 生態学と崩壊:イースター島の事例
生態学と崩壊:イースター島の事例 フラクタル:自己組織化された普遍的構造
フラクタル:自己組織化された普遍的構造