
ヒマワリが太陽の方を向くのは、他の物理システムと同じように、物理法則に従っているからです。 生命力や外部の意図は必要ありません:通常の力学的・生物学的制約だけで十分です。
ラグランジアン \( L = T - V \) は、2つの項、運動エネルギー \(T\) と位置エネルギー \(V\) を用いて、物理システムの進化を記述します。 ヒマワリに適用すると、茎を下に引っ張る重力と、茎を上に曲げる光による成長勾配を比較します。 物理学だけで、残りのすべてを説明できます。
ヒマワリは単に太陽を追っているように見えますが、その運動は微妙なエネルギーバランスの結果です。 18世紀末に ジョゼフ・ルイ・ラグランジュ (1736-1813) によって考案された ラグランジアン は、システムの動的状態を要約する数学的関数です。 具体的には、任意の物体に対して、ラグランジアンは運動エネルギー(運動のエネルギー)と位置エネルギー(蓄えられたエネルギー、例えば重力)の差です。 ただし注意が必要です:ヒマワリの場合、光からのエネルギーは直接運動エネルギーではありません。 それは 静電エネルギー です:光子が細胞に当たると、プロトンポンプを活性化し、膜の両側の電荷を変化させ、水圧を生み出して茎を変形させます。
したがって、ヒマワリのラグランジアンは次のように理解できます:下に引っ張る力(重力、位置エネルギー)と上や光の方に押す力(光子による静電エネルギー)を比較します。 静電エネルギーが優勢な場合、花はまっすぐに立ち、太陽の方を向きます。 重力が優勢な場合(夜や花が老化した時)、植物は傾きます。 ラグランジアンはこれらのエネルギーのいずれでもありません:それらの 差 です。 そして、この差が運動を決定します。
重力 は、すべての質量に作用する力です。 植物が高くて重いほど、その 重力位置エネルギー は大きくなります。 ヒマワリの場合、花の頭(頭状花序)は1キログラム以上の重さがあり、地面から2メートルの高さにあることもあります。 蓄えられた位置エネルギーの量は相当なものです。 このエネルギーは植物内に蓄えられ、茎が曲がると機械的に解放されます。 他の力がなければ、花は必然的に倒れてしまいます。 重力は常に下に引っ張り、あたかもヒマワリを水平に、地面に寝かせようとするかのようです。
植物は毎日、光 という形でエネルギーの流れを受け取ります。 この光は直接的な機械的力ではありません:放射エネルギーを運ぶ光子です。 しかし、植物細胞の内部では、このエネルギーが正確な生物学的反応の連鎖を引き起こします。
光子はまず、茎の光が当たる側にある フォトトロピン というタンパク質を活性化します。 これらのタンパク質は、成長ホルモンである オーキシン を陰の側に再分配します。 オーキシンは到達した細胞の壁を酸性化し、セルロース繊維を緩め、これらの細胞がさらに伸びることを可能にします。 そのため、陰の側は光が当たる側よりも速く伸び、茎は機械的に光の方に曲がります。
この過程には意図は介在しません。 局所的な化学的違いだけで、方向性のある運動を生み出すことができ、これは化学と材料力学の法則を適用するだけで実現します。
ヒマワリの運動は、全体的な静電エネルギーによるものではなく、光によって誘発される植物組織内の圧力の不均衡によるものです。 この現象は 向光性 と呼ばれ、イオン勾配 (\(K^+\), \(Ca^{2+}\), \(H^+\)) によって細胞の局所的な電位が変化します。 これらの勾配は、組織内の水の再分配を引き起こし、細胞の内部圧力(膨圧)を変化させます。 膨圧の低い側はより伸びます、なぜならその細胞は膨張に対する機械的抵抗が低いからです。
茎の両側のこの伸びの違いが、徐々に曲がりを作り、植物をより剛性の高い側、つまり全体的に光の方向に向けます。
力学的な観点から見ると、この変形は、茎の曲がりに関連する弾性位置エネルギーと、膨圧に関連する圧力エネルギーのバランスの結果です。 組織の粘性散逸は、進化を遅く、減衰させ、ほとんど慣性なしにします。
したがって、茎は 粘弾性 構造のように振る舞い、その全体的な形は内部制約の継続的な再調整から生まれます。
生物システム「ヒマワリ」は、計算や意識的な最適化を行うことはありません。 その振る舞いは、太陽エネルギーが常に流れ込む 生きているシステム に物理法則を適用した結果にすぎません。
最小作用の原理は、力学において、システムが運動エネルギーと位置エネルギーをバランスさせながら全体的に進化する方法を記述します。これは単純なエネルギーを最小化するのではなく、物理学によって課せられた全体的な制約に従うことです。
生きている植物では、組織は常に水とエネルギーで供給され、その一部を熱と内部抵抗という形で散逸させます。 これらの交換は、茎の剛性と形状を継続的に変化させます。
ヒマワリの成長と向きは、機械的により安定で光に適した構成への徐々な調整として現れます。 この振る舞いは意図的な戦略ではなく、物理法則と進化の直接的な結果です。
ヒマワリの運動に奇跡的なものは何もありません。 それは、物質的構造に常に作用する普通の物理的制約の結果にすぎません:
これら4つの効果はすべて知られており測定可能で、ヒマワリの振る舞いを完全に説明するのに十分です。 生命力、意図、設計者は、この運動を説明するために必要ありません。
ヒマワリが示すことは、すべての生きているシステムに当てはまります:振る舞いの複雑さは謎の証拠ではなく、組織化された物質に適用された単純な物理法則の予測可能な結果です。生命は物理学を免れるものではなく、その一つの表現にすぎません。
ヒマワリは向光性のおかげで太陽を追います。植物ホルモンが茎の非対称的な成長を引き起こし、徐々に花を光の方向に向けます。
ラグランジアンは、運動エネルギーと位置エネルギーの差として定義される数学的関数です: \( L = T - V \) これは、物理システムの最適な動的進化を決定することを可能にします。
はい、例外なく。生きている生物は、重力、熱力学、電磁気学、流体力学など、あらゆる物理システムと同じ法則に従う原子と分子で構成されています。 生命は物理学の例外ではなく、その特に複雑な組織形態です。
いいえ。振る舞いや構造の複雑さは、外部の意図の証拠ではありません:長期間にわたって組織化された物質に適用された単純な物理的・化学的法則の結果です。 ヒマワリは無数の例の一つにすぎません。
向光性は、植物が光に反応して成長の方向を調整する能力です。 これは、茎の細胞の非対称的な伸長を引き起こすホルモン、オーキシンの再分配に基づいています。 これは純粋に化学的・力学的なメカニズムで、光の捕捉を改善するために進化によって選択されたものです—意図は介在しません。
植物は、重力、弾性、水圧、エネルギー散逸などの物理的力にさらされる物質的構造を持っています。したがって、力学のツールを使用して、その振る舞いの特定の側面をモデル化することができます。
いいえ、厳密な意味では。ヒマワリは何も最小化せず、最適を求めることもありません:それは局所的な物理的制約に、瞬間瞬間従っています。 結果が時々変分解に似ているように見える場合でも、それは同じ物理法則が至るところに適用されるからであって、植物が目標を追求したり、外部の意図がその成長を導いたりするからではありません。
はい。変分法やラグランジアンの方法は、生体力学、細胞モデリング、生物流体力学、理論神経科学で使用されています。
 致命的不均衡:極小の物質の予想外の力
致命的不均衡:極小の物質の予想外の力  進化の大フィルター:フェルミのパラドックスの鍵
進化の大フィルター:フェルミのパラドックスの鍵  なぜヒマワリは太陽の方を向くのか?ラグランジアンによる解答
なぜヒマワリは太陽の方を向くのか?ラグランジアンによる解答  世界人口2026:大陸別の人口動向
世界人口2026:大陸別の人口動向  なぜ生命は不均衡から生まれ、熱力学的平衡で消滅するのか
なぜ生命は不均衡から生まれ、熱力学的平衡で消滅するのか  電磁スペクトルと視覚:私たちの目が捉える地球の姿
電磁スペクトルと視覚:私たちの目が捉える地球の姿  自己と非自己:物理法則から見たアイデンティティの簡単な解説
自己と非自己:物理法則から見たアイデンティティの簡単な解説  分子時計:突然変異の偶然性から時間の測定へ
分子時計:突然変異の偶然性から時間の測定へ  ホワイトサンズの足跡:アメリカ大陸の最初の一歩
ホワイトサンズの足跡:アメリカ大陸の最初の一歩  ホミニン:出現、拡散、絶滅
ホミニン:出現、拡散、絶滅 主要な自然災害:最も可能性の高い脅威は何か?
主要な自然災害:最も可能性の高い脅威は何か? 文明の大崩壊:重要な時期と原因
文明の大崩壊:重要な時期と原因 出生率の低下:人口災害か自然な進化か?
出生率の低下:人口災害か自然な進化か? 自然選択 vs 偶然:なぜ進化は宝くじではないのか?
自然選択 vs 偶然:なぜ進化は宝くじではないのか? 生命が地球から始まったら?パンスペルミア理論の革命
生命が地球から始まったら?パンスペルミア理論の革命 世界を激変させる大分岐:生存か崩壊か?
世界を激変させる大分岐:生存か崩壊か? 原始化学:最初の有機分子はどこで生まれたのか?
原始化学:最初の有機分子はどこで生まれたのか? COとCO₂:2つのガス、2つのリスク、2つの生物学的メカニズム
COとCO₂:2つのガス、2つのリスク、2つの生物学的メカニズム 自発的同期:物理学から生命までの普遍的現象
自発的同期:物理学から生命までの普遍的現象 時間的課題:10億年をどのように視覚化するか?
時間的課題:10億年をどのように視覚化するか? 生命の誕生に不可欠な3つの要素
生命の誕生に不可欠な3つの要素 なぜホモ属は90万年前に絶滅の危機に瀕したのか?
なぜホモ属は90万年前に絶滅の危機に瀕したのか? 影の生物圏
影の生物圏 人間中心主義の衰退
人間中心主義の衰退 カブトガニ:生きている化石!
カブトガニ:生きている化石! 宇宙における生命の存在:バイオシグネチャー
宇宙における生命の存在:バイオシグネチャー 人工知能の課題と脅威
人工知能の課題と脅威 生命の起源:パンスペルミア理論
生命の起源:パンスペルミア理論 生命の起源:ホワイトスモーカー理論
生命の起源:ホワイトスモーカー理論 なぜ37度セルシウスなのか?
なぜ37度セルシウスなのか? 私たちは宇宙で孤独なのか?科学と推測の間で
私たちは宇宙で孤独なのか?科学と推測の間で 氷の中の生命の痕跡:先史時代のマムートの出現
氷の中の生命の痕跡:先史時代のマムートの出現 ドリアス期:メガファウナを絶滅させたミニ氷河期
ドリアス期:メガファウナを絶滅させたミニ氷河期 2つの大氷河期:凍った地球の海で生き残る
2つの大氷河期:凍った地球の海で生き残る 動物の切断後の再生:器官の再生
動物の切断後の再生:器官の再生 生命の果て:地底のメフィスト、深淵の虫
生命の果て:地底のメフィスト、深淵の虫 宇宙で固体フラーレンが発見される
宇宙で固体フラーレンが発見される 人間の歩行:原人の二足歩行の起源
人間の歩行:原人の二足歩行の起源 カラボ:人間進化の窓
カラボ:人間進化の窓 エントロピー:時間とは何か?
エントロピー:時間とは何か? 無生物から生命への移行
無生物から生命への移行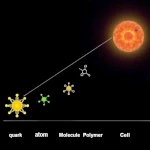 複雑さの物語:素粒子から最初の生物まで
複雑さの物語:素粒子から最初の生物まで メガポード:火山の熱を利用する
メガポード:火山の熱を利用する アルディピテクス:440万年前のエチオピアの原人
アルディピテクス:440万年前のエチオピアの原人 自然選択:カバマダラの例
自然選択:カバマダラの例 オルドビス紀:サンゴ、三葉虫、放散虫の時代
オルドビス紀:サンゴ、三葉虫、放散虫の時代 液体の水:単なる溶媒以上、化学反応の促進剤
液体の水:単なる溶媒以上、化学反応の促進剤 ネアンデルタール人:人類の失われたいとこ
ネアンデルタール人:人類の失われたいとこ アシモ:未来のヒューマノイド
アシモ:未来のヒューマノイド 生命の誕生を可能にした条件は何か?
生命の誕生を可能にした条件は何か? フェルミのパラドックスとプラトンの洞窟:私たちは孤独か、それとも盲目か?
フェルミのパラドックスとプラトンの洞窟:私たちは孤独か、それとも盲目か? クマムシ:生物学の法則に挑戦する不死身の生物
クマムシ:生物学の法則に挑戦する不死身の生物 トゥーマイ:最古の原人の一つ
トゥーマイ:最古の原人の一つ 生命の樹:数十億の絶滅種と単一の祖先コミュニティ
生命の樹:数十億の絶滅種と単一の祖先コミュニティ 深海の生命:極限の適応を遂げた生物
深海の生命:極限の適応を遂げた生物 シアノバクテリアと酸素危機:原始的な環境災害
シアノバクテリアと酸素危機:原始的な環境災害 物質から生命へ:生物学的出現の曖昧な境界
物質から生命へ:生物学的出現の曖昧な境界 世界最小のカエル:微小脊椎動物の生理的秘密
世界最小のカエル:微小脊椎動物の生理的秘密 小氷期の説明
小氷期の説明 生命の光:月が明かすバイオシグネチャー
生命の光:月が明かすバイオシグネチャー 生きている光:生物発光の驚異的な秘密
生きている光:生物発光の驚異的な秘密 感覚を超えて:科学の大革命
感覚を超えて:科学の大革命 原始のスープ:地球生命の化学的揺籃
原始のスープ:地球生命の化学的揺籃 世界人口:10億人から人口飽和へ
世界人口:10億人から人口飽和へ 生態学と崩壊:イースター島の事例
生態学と崩壊:イースター島の事例 フラクタル:自己組織化された普遍的構造
フラクタル:自己組織化された普遍的構造