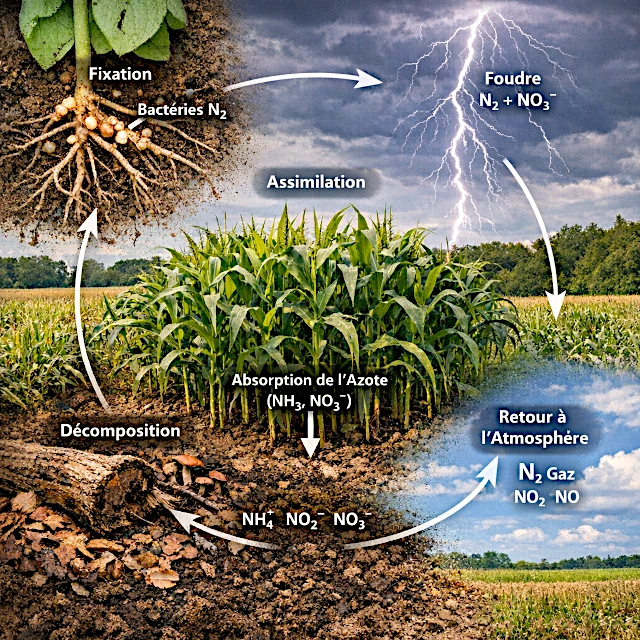
窒素はタンパク質やDNAを作るために欠かせない元素ですが、大気中に豊富に存在する気体のままでは多くの生物が利用できません。窒素循環は、細菌、植物、動物、そして自然の働きによって、この窒素を生物が利用できる形へと絶えず変換しています。これらの変換がなければ、土壌の肥沃度は失われ、海洋の生産性は低下し、生態系は崩壊してしまいます。この目に見えないが極めて重要な循環が、地球の生物学的バランスを支えています。
地球の大気には、体積で約78%、質量で75%の窒素ガス(\(N_2\))が含まれています。 しかしながら、この巨大な潜在的資源は、ほとんどの生物にとって利用不可能です。 動物、人間、植物のいずれも、これを栄養素に変換することはできません。
その理由は、2つの窒素原子(N≡N)の間の三重結合の例外的な強度にあります。 この結合を切断するには、莫大なエネルギーが必要です。
2つの経路があります:30,000°Cの雷、または細菌の化学的な優雅さとそのニトロゲナーゼ酵素で、\(N_2\)を\(NH_3\)(窒素ガス→アンモニア)に還元できます。 この事前の切断がなければ、タンパク質、DNA、筋肉、神経、血液は存在しません:生命は存在しません。
窒素循環は、不活性ガスを同化可能な栄養素に変換し、その後大気に戻す一連の変換です。
N.B.:アントワーヌ・ラヴォアジエ(1743-1794)は、「窒素」(ギリシャ語の「a-zôê」、生命なし)という用語を提案しました。 皮肉なことに、このガス状態では不活性な元素が、固定されるとすべての生細胞に不可欠であることが明らかになります。
| プロセス | 化学変換 | 細菌 + 菌類 | 必要条件 | 生態学的役割 |
|---|---|---|---|---|
| 共生固定 | \(N_2 \rightarrow NH_3\) | Rhizobium, Bradyrhizobium | マメ科植物の根との共生、微好気性 | 栽培植物への直接的な窒素供給 |
| 自由固定 | \(N_2 \rightarrow NH_3\) | Azotobacter(好気性)、Clostridium(嫌気性) | 土壌、水環境、利用可能な有機炭素 | 自然の肥沃性への拡散的だが持続的な貢献 |
| 硝化(ステップ1) | \(NH_3 \rightarrow NO_2^-\) | Nitrosomonas, Nitrosospira | 厳格な好気性、中性から弱アルカリ性のpH | 亜硝酸塩の形成、硝酸塩の前駆体 |
| 硝化(ステップ2) | \(NO_2^- \rightarrow NO_3^-\) | Nitrobacter, Nitrospira | 好気性、広い温度範囲への耐性 | 硝酸塩の生産、植物にとって高度に同化可能な形態 |
| 脱窒 | \(NO_3^- \rightarrow N_2\)(\(NO_2^-\), NO, \(N_2O\)経由) | Pseudomonas, Paracoccus, Bacillus | 無酸素(飽和土壌、堆積物)、不安定な有機物質 | 循環の閉鎖、\(N_2\)の大気への戻り |
| アンモニフィケーション(鉱質化) | 有機N \(\rightarrow NH_4^+\) | 腐生菌(Bacillus, Streptomycesなど) | 好気性または嫌気性、死体の分解 | 有機廃棄物からの窒素のリサイクル |
N.B.:人間活動による合成肥料は、1950年以来、陸上生態系に入る反応性窒素の流れを2倍に増加させました。
産業革命以前、生物学的および大気的固定の自然な流れは、土壌や水中の反応性窒素の安定した在庫を維持していました。 ユストゥス・フォン・リービッヒ(1803-1873)の研究は、窒素化合物の肥沃性への重要性を既に予見していました。 しかし、ハーバー・ボッシュ法(20世紀初頭)の発明が、合成窒素肥料の生産を飛躍的に増加させました。 現在、人為的固定は自然固定を上回っています。
過剰な硝酸塩は雨によって流出し、川を経て沿岸域に達し、富栄養化したデッドゾーンを引き起こします。ここで藻類の大量発生が水生動物を窒息させます。 メキシコ湾やバルト海は、この拡散汚染を例示しています。 さらに、硝化および脱窒の副産物である一酸化二窒素(\(N_2O\))は、二酸化炭素の300倍の温室効果を持ち、地球温暖化に寄与しています。
大気中の窒素を生物が利用できる形に変換し、再び大気へ戻す一連のプロセスのことです。
N₂分子は非常に安定しており、分解できるのは特定の細菌だけだからです。
窒素固定、硝化、アンモニア化、脱窒といった主要な段階を担っています。
土壌中の硝酸塩やアンモニウムを吸収し、タンパク質やアミノ酸、DNAを合成します。
肥料、汚染、産業排出により反応性窒素が過剰となり、富栄養化や生態系の乱れを引き起こします。
 最小作用の原理:なぜ自然は常に最も経済的な道を選ぶのか?
最小作用の原理:なぜ自然は常に最も経済的な道を選ぶのか?  収斂進化:偶然、必然…それとも幻?
収斂進化:偶然、必然…それとも幻?  五大大量絶滅:これらの地球規模の災厄は何を語るのか?
五大大量絶滅:これらの地球規模の災厄は何を語るのか?  氷圏:氷の巨人がまだ私たちを守っている
氷圏:氷の巨人がまだ私たちを守っている  窒素循環:空気から生命へ
窒素循環:空気から生命へ  ジェット気流:大気の川が氾濫するとき
ジェット気流:大気の川が氾濫するとき  炭素サイクル:海洋、森林、地下の調和的な相互作用
炭素サイクル:海洋、森林、地下の調和的な相互作用  水のパラドックス:水なしに生命なし
水のパラドックス:水なしに生命なし  なぜ220V交流がコンセントに?物理学の問題
なぜ220V交流がコンセントに?物理学の問題  最後の氷河期ショック:ヤンガードライアスと先史時代の社会への影響
最後の氷河期ショック:ヤンガードライアスと先史時代の社会への影響  536年:夏のない年と後期古代小氷期の始まり
536年:夏のない年と後期古代小氷期の始まり  最小細胞:避けられない出現
最小細胞:避けられない出現 

 世界のエネルギー消費の進化:蒸気機関からデジタルサーバーまで
世界のエネルギー消費の進化:蒸気機関からデジタルサーバーまで  人口転換:成長か衰退か?
人口転換:成長か衰退か?  アガシーズ湖:気候を変えた大洪水
アガシーズ湖:気候を変えた大洪水  酔っ払いの森:永久凍土の融解現象を理解する
酔っ払いの森:永久凍土の融解現象を理解する  空気中の電気!
空気中の電気!  なぜCO2は地面に落ちないのか?
なぜCO2は地面に落ちないのか?  エルニーニョとラニーニャ:太平洋の二つの息吹が地球の気候を律する
エルニーニョとラニーニャ:太平洋の二つの息吹が地球の気候を律する  絶滅危惧種:絶滅のダイナミクス
絶滅危惧種:絶滅のダイナミクス  酸性雨の有毒な遺産
酸性雨の有毒な遺産  気候変動の数字:科学データが語るもの
気候変動の数字:科学データが語るもの  持続可能な開発とは何か?
持続可能な開発とは何か?  アラル海:人類への教訓
アラル海:人類への教訓  三峡ダム:生態系への影響
三峡ダム:生態系への影響  観測史上最も暑い年
観測史上最も暑い年  今後の気候変動の規模
今後の気候変動の規模  危機に瀕する地球:崩壊か再生か?
危機に瀕する地球:崩壊か再生か?  危機に瀕する氷:北極海氷の避けられない減少
危機に瀕する氷:北極海氷の避けられない減少  地球の水資源:海洋から地下水まで
地球の水資源:海洋から地下水まで  海面上昇の速度はどれくらいか?
海面上昇の速度はどれくらいか?  もう一つの気候の脅威:地球が影と湿気に包まれる
もう一つの気候の脅威:地球が影と湿気に包まれる  サハラの年齢:緑の草原から石の砂漠へ
サハラの年齢:緑の草原から石の砂漠へ  1800年から2100年までの世界人口
1800年から2100年までの世界人口  石油:再生可能エネルギー時代における巨人の没落
石油:再生可能エネルギー時代における巨人の没落  カムチャツカのタラバガニ:海の巨人
カムチャツカのタラバガニ:海の巨人  イースター島:社会の崩壊
イースター島:社会の崩壊  世界のエコロジカル・フットプリントのわずかな進展
世界のエコロジカル・フットプリントのわずかな進展  南大西洋異常
南大西洋異常  地盤沈下現象
地盤沈下現象  なぜ地球はこんなに頻繁に曇っているのか?
なぜ地球はこんなに頻繁に曇っているのか?  地球の新しい雲
地球の新しい雲 デザーテック:ヨーロッパとアフリカのための太陽エネルギーの夢
デザーテック:ヨーロッパとアフリカのための太陽エネルギーの夢