
進化は同じ制約に対して同じ解をもたらすからです:生物が似た環境で生活したり、類似した課題に直面したりすると、自然選択はそれらを類似した形、機能、または行動に向かわせます。最近の共通の祖先がいなくてもです。これが収斂進化の原理です。この原理が、イルカと魚竜が流体力学的な体型を共有する理由、タコの目が脊椎動物の目に似ている理由、砂漠の植物が同じ生存戦略を発達させる理由を説明します。自然は無数の道を探求しますが、物理と生物の法則はしばしば解を同じ構造に導きます。
収斂進化は、進化の中で最も魅力的な現象の一つです。 数億年にわたって分かれた非常に遠い系統の種が、驚くほど似た形質を発達させることがあります。 これらの類似性は、別々の進化の結果として提示されていますが、本当にそのメカニズムだけでこれほど似た解決策を説明できるのか、疑問が残ります。
捕食者から逃れるために速く泳ぐ、深海で光を感知する、空を征服する、温度変化に耐える、 低酸素環境でのガス交換を最適化する、極限の圧力に耐える、不足時にエネルギーを節約する、化学信号を検出する、 または捕食を避けるために擬態する:これらはすべて環境によって課せられた制約です。
理論によれば、進化のメカニズムは、計画も目的も意図も持たず、世代を重ねるごとに最も効果的な解決策を探索し選択します。 しかし、非常に遠い系統が同じ答えにたどり着くことを繰り返すとき、疑問が生じます:自然は本当に再発明しているのか、それとも私たちが考える以上に決まった道をたどっているのか? あたかも進化が、ある種の発明を繰り返すように、ほとんどやむを得ずに押し進められているかのようです。
この現象は、完全に偶然的で偶発的な進化という考えに挑戦しているようです。 もはや偶然だけではなく、ある種の繰り返しの傾向があるのかもしれません:自然選択は、柔軟性のない技術者のように、種を限られた効果的な解決策のセットに押しやります。 あたかも物質とエネルギーの法則が、背景で生命の可能な形態を強制しているかのようです。 飛ぶための翼、見るための目、獲物を無力化するための毒…これらは直接的な関連のない系統で現れた革新であり、自然が私たちがかろうじて垣間見る指示書に従っているかのようです。
進化を主に予測不可能で偶発的なプロセスと見なした古生物学者スティーブン・ジェイ・グールド(1941–2002)は、収斂が不安な疑問を投げかけると認めていました: もしある生命の形態が偶然の産物だけでなく、より深く、もしかすると避けられない制約の表現であるとしたらどうでしょう? 彼のエッセイ『Wonderful Life』では、「生命のテープを再生する」ことを想像し、こう問いました:進化の歴史が再び始まったら、同じ生物、あるいは存在の課題に対する同じ解決策が再び現れるでしょうか? 収斂は繰り返しを示唆していますが、進化の中で本当に偶然なのか、疑問が残ります。
この考えは、目まいがするような視点を開きます:生命の多様性は、その見かけの豊かさにもかかわらず、私たちが想像する以上に予測可能な枠組みに収まるのかもしれません。 物理的世界の制約がフィルターとして働き、ある種の革新だけを浮上させるのです—それらは、奇妙なことに、非常に遠い起源を持つ種に現れることになります。 謙虚さの教訓かもしれませんが、同時に、自然の創造性を導く、まだ十分に理解されていないより深いルールが存在するのではないかと考えさせられます。
| 収斂的とされる形質 | 独立したとされるグループ | 適応的機能 | 分岐の年代 | 例と特徴 |
|---|---|---|---|---|
| 単一レンズの目 | 脊椎動物 / 頭足類 | 像形成 | > 5.5億年前 | 人間(盲点あり) / イカ(盲点なし)——遺伝的遺産を共有しながら驚くべき類似性 |
| 能動的飛行(翼) | 昆虫 / 翼竜 / 鳥 / コウモリ | 空中移動 | > 3.5億年前 | 4つの異なる体制計画、しかし空力学的制約が似た解決策を強制しているように見える |
| カニ化(カニの形) | 本物のカニ / 偽カニ(異尾下目) | 保護、横歩き | > 2億年前 | 「カニ化」:遠い系統が同じシルエットを採用、おそらく似た制約の下で |
| 反響定位 | コウモリ / イルカ / アマツバメ | 暗闇でのナビゲーションと狩り | > 8000万年前 | 似た音響メカニズム、しかし時折驚くほど収斂的な遺伝的基盤 |
| 擬態(同色性) | 昆虫 / 鳥 / 哺乳類 | 隠蔽 | > 3億年前 | ナナフシ、ランフラワーマント、ホッキョクウサギ——似た戦略、しかし必ずしも独立した生化学的経路ではない |
| 麻痺性の毒 | ヘビ / ヤモリ / クモ / サソリ / カエル | 獲物の不動化または防御 | > 2億年前 | 似た毒素、時折共通の祖先に既に存在した遺伝子ファミリーに基づく |
| 生物発光 | 昆虫 / 深海魚 / 細菌 / キノコ / イカ | 誘引、おとり、擬態 | > 5億年前 | ルシフェリンとルシフェラーゼ:同じ化学反応、しかし本当に独立した起源は議論の余地あり |
| 極限環境耐性(放射線耐性) | 古細菌 / 細菌 / クマムシ | 極限の耐性 | > 27億年前 | DNA修復メカニズムの類似性、おそらく非常に古い分子モジュールに由来 |
収斂進化のすべての例の中で、目はおそらく最も壮観であり、同時に最も当惑させられるものです。 生物学者がその起源を探るほど、単純で明確な収斂という考えは複雑になっていきます。
タコと人間の共通の祖先は、約5億5000万から6億年前(後期先カンブリア時代)に生息していた原始的な両側対称動物でした。 この動物は、水晶体、網膜、瞳孔を持つ構造化された目を持っていませんでしたが、視覚装置を全く持っていなかったわけではありません: 既に原始的な光受容体、Pax6遺伝子、 目の発達の真の指揮者であり、また祖先的オプシンを動物系統全体で共有していました。
まさにここで、収斂の概念はそれほど明確ではなくなります。 1983年、Walter Jakob Gehring(1939–2014)が、マウスのPax6遺伝子をショウジョウバエに注入すると触角に目が形成されることを示したとき、 彼は、目の基礎となる遺伝子プログラムが数億年にわたって分かれた動物に共通していることを明らかにしました。 タコと人間は「ゼロから目を再発明」したのではなく、それぞれの方法で同じ祖先的な分子キットを再活性化し、驚くほど似た構造に至ったようです。
退化とは、現代の生物に数百万年前に消失した古代の形質が再び現れることを指します。 この現象は、進化が形質を本当に「再発明」しているわけではないことを示しています:祖先的な遺伝子プログラムがゲノム内にまだ存在し、沈黙しているだけかもしれません。 歯を持つニワトリ、足を持つクジラ、または四肢の原基を示すヘビは、生命が古代の構造を再活性化する能力を示しています。
退化は収斂進化の解釈を複雑にします:独立した発明として提示されるものが、既存の遺伝子モジュールの再活性化に過ぎないかもしれません。 言い換えれば、一部の「収斂」は、環境の制約によって課せられた新しい解決策ではなく、生命の深い歴史に刻まれた古代の状態への可能な回帰かもしれません。
チャールズ・ダーウィン(1809–1882)が1859年に提案した枠組みは、3つの主要な原則に基づいています:共通の祖先からの修飾を伴う子孫、個体間の遺伝的変異、そして自然選択としての選別メカニズム。 これら3つの柱は、現在、遺伝学、古生物学、分子系統学、直接観察によって確認されています。 科学コミュニティの真面目なメンバーは誰もこれらを疑っていません。
ダーウィンは本質を捉えていましたが、彼が提案したメカニズムだけではすべてを説明するには不十分です。 21世紀の生物学は彼の業績に矛盾しません:それを拡張し、補完し、豊かにし、時には想像を超えています。
収斂進化は、環境や物理法則が進化を偶然だけに任せず、似た解決策に導くことができることを示しています。 これにより、類似の制約が働く場所ならどこでも、地球外知的生命体が出現する可能性——確実性ではない——が開かれます。 目、翼、または類似の社会構造が現れるかもしれません…または、生命が私たちが想像する以上に決まった道をたどることを示唆するかもしれません。
しかし、この理論的可能性は、より深い現実を覆い隠してはなりません:進化の軌跡は、依然としてユニークで予測不可能な出来事の連鎖によって形作られています。
それでも、これらの不思議な類似性にもかかわらず、地球上の生命は並外れて偶発的な歴史の産物であり、 不可能で再現不可能な出来事の連続によって形作られています。 この道のりが、その断絶、事故、迂回とともに、他の場所で再現されるという兆候はありません;逆に、それが地球とそのユニークな歴史に固有のものであることを示唆しています。
収斂進化とは、近い共通祖先を持たない生物種が、独立に似た特徴を獲得する現象です。これは、同様の環境圧が働き、進化が類似した解決策へと導かれるために起こります。
いいえ。収斂進化はあらかじめ決められた計画ではありません。物理的・生物学的・生態学的な制約が、取り得る解の範囲を限定することで生じます。進化は偶然性を含みますが、無制限ではありません。
同じ問題を解決するために特に効率的な形態が存在するためです。例えば、速く泳ぐ、飛ぶ、暗闇で見る、圧力に耐えるなど。環境は最も効果的な解を選択するフィルターとして働きます。
ある程度の予測可能性を示唆します。同じ制約下では、生物は似た構造へ進化しやすくなります。しかし生命の多様性は、多くの進化経路が依然として存在することを示しています。
鳥類・コウモリ・昆虫の翼、頭足類と脊椎動物の眼、魚類・イルカ・イクチオサウルスの流線型の体、アリとシロアリの社会戦略などです。
はい。頭足類、カラス類、霊長類、海生哺乳類など、複数の系統が独立に高度な認知能力を発達させています。これは、適切な生態条件があれば知性が生じ得ることを示唆します。
有効なヒューリスティック(探索指針)となります。普遍的に効率の良い解があるなら、同様の制約下にある他の惑星でも再び現れる可能性があります。これは宇宙生物学の仮説形成に役立ちます。
いいえ。むしろ自然な結果です。自然選択は、物理・化学・生物の法則によって制限された可能性空間を探索します。収斂進化はこれらの制約の強さを示しています。
 最小作用の原理:なぜ自然は常に最も経済的な道を選ぶのか?
最小作用の原理:なぜ自然は常に最も経済的な道を選ぶのか?  収斂進化:偶然、必然…それとも幻?
収斂進化:偶然、必然…それとも幻?  五大大量絶滅:これらの地球規模の災厄は何を語るのか?
五大大量絶滅:これらの地球規模の災厄は何を語るのか?  氷圏:氷の巨人がまだ私たちを守っている
氷圏:氷の巨人がまだ私たちを守っている  窒素循環:空気から生命へ
窒素循環:空気から生命へ  ジェット気流:大気の川が氾濫するとき
ジェット気流:大気の川が氾濫するとき  炭素サイクル:海洋、森林、地下の調和的な相互作用
炭素サイクル:海洋、森林、地下の調和的な相互作用  水のパラドックス:水なしに生命なし
水のパラドックス:水なしに生命なし  なぜ220V交流がコンセントに?物理学の問題
なぜ220V交流がコンセントに?物理学の問題  最後の氷河期ショック:ヤンガードライアスと先史時代の社会への影響
最後の氷河期ショック:ヤンガードライアスと先史時代の社会への影響  536年:夏のない年と後期古代小氷期の始まり
536年:夏のない年と後期古代小氷期の始まり  最小細胞:避けられない出現
最小細胞:避けられない出現 

 世界のエネルギー消費の進化:蒸気機関からデジタルサーバーまで
世界のエネルギー消費の進化:蒸気機関からデジタルサーバーまで  人口転換:成長か衰退か?
人口転換:成長か衰退か? 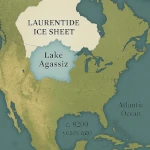 アガシーズ湖:気候を変えた大洪水
アガシーズ湖:気候を変えた大洪水  酔っ払いの森:永久凍土の融解現象を理解する
酔っ払いの森:永久凍土の融解現象を理解する  空気中の電気!
空気中の電気! 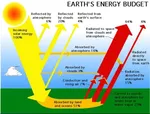 なぜCO2は地面に落ちないのか?
なぜCO2は地面に落ちないのか?  エルニーニョとラニーニャ:太平洋の二つの息吹が地球の気候を律する
エルニーニョとラニーニャ:太平洋の二つの息吹が地球の気候を律する  絶滅危惧種:絶滅のダイナミクス
絶滅危惧種:絶滅のダイナミクス  酸性雨の有毒な遺産
酸性雨の有毒な遺産  気候変動の数字:科学データが語るもの
気候変動の数字:科学データが語るもの  持続可能な開発とは何か?
持続可能な開発とは何か?  アラル海:人類への教訓
アラル海:人類への教訓  三峡ダム:生態系への影響
三峡ダム:生態系への影響 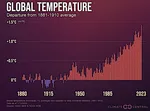 観測史上最も暑い年
観測史上最も暑い年  今後の気候変動の規模
今後の気候変動の規模  危機に瀕する地球:崩壊か再生か?
危機に瀕する地球:崩壊か再生か?  危機に瀕する氷:北極海氷の避けられない減少
危機に瀕する氷:北極海氷の避けられない減少  地球の水資源:海洋から地下水まで
地球の水資源:海洋から地下水まで  海面上昇の速度はどれくらいか?
海面上昇の速度はどれくらいか?  もう一つの気候の脅威:地球が影と湿気に包まれる
もう一つの気候の脅威:地球が影と湿気に包まれる  サハラの年齢:緑の草原から石の砂漠へ
サハラの年齢:緑の草原から石の砂漠へ  1800年から2100年までの世界人口
1800年から2100年までの世界人口  石油:再生可能エネルギー時代における巨人の没落
石油:再生可能エネルギー時代における巨人の没落  カムチャツカのタラバガニ:海の巨人
カムチャツカのタラバガニ:海の巨人  イースター島:社会の崩壊
イースター島:社会の崩壊  世界のエコロジカル・フットプリントのわずかな進展
世界のエコロジカル・フットプリントのわずかな進展  南大西洋異常
南大西洋異常  地盤沈下現象
地盤沈下現象  なぜ地球はこんなに頻繁に曇っているのか?
なぜ地球はこんなに頻繁に曇っているのか?  地球の新しい雲
地球の新しい雲 デザーテック:ヨーロッパとアフリカのための太陽エネルギーの夢
デザーテック:ヨーロッパとアフリカのための太陽エネルギーの夢