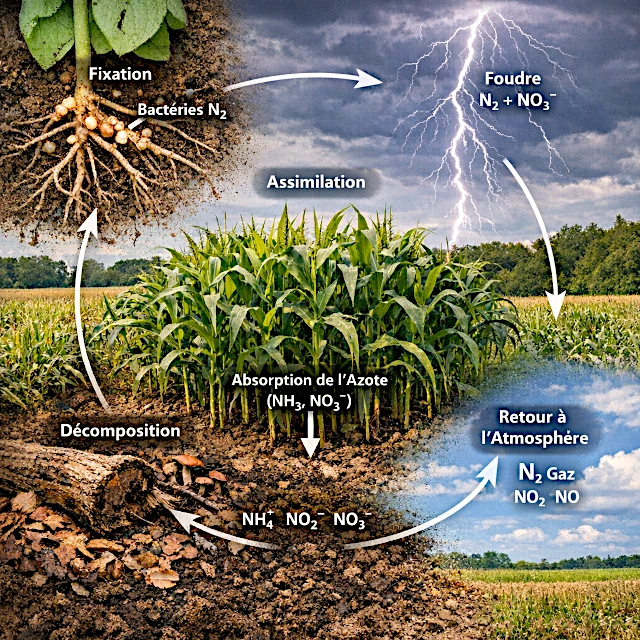
由于构建蛋白质和DNA所必需的氮元素,在大气中虽以气态形式大量存在,却无法被大多数生物直接利用。氮循环通过细菌、植物、动物及自然现象的协同作用,持续将这种元素转化为生物可吸收的形态。若没有这些连续转化过程——固氮、硝化、同化、氨化及反硝化——生态系统将迅速崩溃。这一无形却至关重要的循环维系着地球的生态平衡,决定着土壤肥力、海洋生产力及气候稳定性。
地球大气中,按体积计算,氮气(\(N_2\))含量接近78%,按质量计算则为75%。然而,这一对生命而言潜力巨大的资源,对大多数生物来说却无法利用。没有任何动物、人类或植物能够将其转化为营养物质。
原因在于两个氮原子之间的三重化学键(N≡N)强度极高,需要相当大的能量才能打破这个键。
存在两条路径:3万摄氏度下的闪电,或细菌及其固氮酶所展现的化学精妙——后者能将\(N_2\)还原为\(NH_3\)(二氮→氨)。若无此先决裂解,便无蛋白质、无DNA、无肌肉、无神经元、无血液:亦无生命。
氮循环是一系列转化过程,将惰性气体转化为可吸收的营养物质,随后再将其释放回大气中。
注:安托万·拉瓦锡(1743-1794)提出了“氮”这一术语(源自希腊语“a-zôê”,意为“无生命”)。矛盾的是,这种气态惰性元素一旦被固定,便成为每个活细胞不可或缺的物质。
| 过程 | 化学转化 | 细菌 + 真菌 | 必要条件 | 生态角色 |
|---|---|---|---|---|
| 共生固氮 | \(N_2 \rightarrow NH_3\) | 根瘤菌属,慢生根瘤菌属 | 与豆科植物根部的共生,微需氧性 | 直接向栽培植物供应氮素 |
| 自由固定 | \(N_2 \rightarrow NH_3\) | 固氮菌属(好氧)、梭菌属(厌氧) | 土壤、水生环境、可利用有机碳 | 广泛但持续的自然肥力贡献 |
| 硝化作用(第一步) | \(NH_3 \rightarrow NO_2^-\) | 亚硝化单胞菌属、亚硝化螺菌属 | 严格需氧,中性至微碱性pH | 亚硝酸盐的形成,硝酸盐的前体 |
| 硝化作用(第二步) | \(NO_2^- \rightarrow NO_3^-\) | 硝化杆菌属,硝化螺菌属 | 好氧性,对广泛温度范围的耐受性 | 硝酸盐的生产,植物高度可吸收的形式 |
| 反硝化作用 | \(NO_3^- \rightarrow N_2\)(经由 \(NO_2^-\)、NO、\(N_2O\)) | 假单胞菌属、副球菌属、芽孢杆菌属 | 缺氧(饱和土壤、沉积物),不稳定性有机质 | 循环闭合,\(N_2\) 返回大气 |
| 氨化作用(矿化作用) | 有机氮 \(\rightarrow NH_4^+\) | 腐生真菌(芽孢杆菌、链霉菌等) | 好氧或厌氧条件下,死生物质的分解 | 有机废物中氮的回收利用 |
注:自1950年以来,人类活动通过合成肥料使进入陆地生态系统的活性氮流量翻了一番。
在工业时代之前,生物固氮和大气固氮的自然通量维持了土壤和水中活性氮储量的稳定。尤斯图斯·冯·李比希(1803-1873)的研究早已预见到氮化合物对土壤肥力的重要性。但真正使合成氮肥产量成倍增长的是哈伯-博世法的发明(20世纪初)。如今,人为固氮量已超过自然固氮量。
过量的硝酸盐被雨水淋溶后进入河流,最终汇入沿海区域,导致富营养化的死亡区——藻类大量繁殖使水生动物窒息。墨西哥湾和波罗的海就是这种面源污染的典型例证。此外,一氧化二氮(\(N_2O\))作为硝化与反硝化作用的副产物,其温室效应是二氧化碳的300倍,加剧了全球变暖。
这是将大气中的氮转化为生物可利用形式并最终将其返回大气的一系列过程。
由于N₂分子极其稳定,只有特定细菌才能将其分解。
它们驱动关键步骤:固氮作用、硝化作用、氨化作用和反硝化作用,使氮得以利用或将其返回空气中。
它们从土壤中吸收硝酸盐和铵,以构建蛋白质、氨基酸和DNA。
化肥、污染和工业排放增加了过量的活性氮,导致富营养化、生态失衡和温室气体释放。
 最小作用量原理:大自然为何总是选择最经济的路径?
最小作用量原理:大自然为何总是选择最经济的路径?  演化趋同:偶然、必然……还是幻象?
演化趋同:偶然、必然……还是幻象?  五大灭绝事件:这些全球性灾难揭示了什么?
五大灭绝事件:这些全球性灾难揭示了什么?  冰冻圈:当冰之巨人仍在保护我们
冰冻圈:当冰之巨人仍在保护我们 氮循环:从空气到生命
氮循环:从空气到生命  急流:当大气之河决堤
急流:当大气之河决堤  碳循环:海洋、森林与地下土壤的和谐互动
碳循环:海洋、森林与地下土壤的和谐互动  水悖论:没有它,就没有生命
水悖论:没有它,就没有生命  为什么插座里是220V交流电?一个物理学问题
为什么插座里是220V交流电?一个物理学问题  最后一次冰川冲击:新仙女木期及其对史前社会的影响
最后一次冰川冲击:新仙女木期及其对史前社会的影响  536年:没有夏天的一年与晚古小冰期的开始
536年:没有夏天的一年与晚古小冰期的开始  最小细胞:不可避免的涌现
最小细胞:不可避免的涌现 

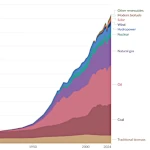 全球能源消费演变:从蒸汽机到数字服务器
全球能源消费演变:从蒸汽机到数字服务器  人口转型:增长还是衰退?
人口转型:增长还是衰退? 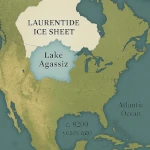 阿加西湖:改变气候的洪水
阿加西湖:改变气候的洪水  醉林:理解永久冻土融化的现象
醉林:理解永久冻土融化的现象  空气中有电!
空气中有电! 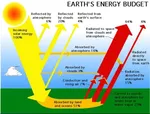 为什么二氧化碳不会掉落到地面?
为什么二氧化碳不会掉落到地面?  厄尔尼诺与拉尼娜:塑造全球气候的太平洋两股呼吸
厄尔尼诺与拉尼娜:塑造全球气候的太平洋两股呼吸  濒危物种:灭绝动力学
濒危物种:灭绝动力学  酸雨的有毒遗产
酸雨的有毒遗产  全球变暖数据:科学数据告诉我们什么
全球变暖数据:科学数据告诉我们什么  什么是可持续发展?
什么是可持续发展?  咸海:给人类的一课
咸海:给人类的一课  三峡大坝:对生态系统的影响
三峡大坝:对生态系统的影响 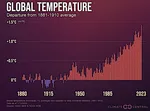 有记录以来最热的年份
有记录以来最热的年份  未来全球变暖的幅度
未来全球变暖的幅度  危机中的地球:崩溃还是复兴?
危机中的地球:崩溃还是复兴?  冰层告急:北极海冰不可阻挡的衰退
冰层告急:北极海冰不可阻挡的衰退  地球的水库:从海洋到地下水
地球的水库:从海洋到地下水  海平面正在上升,但速度有多快?
海平面正在上升,但速度有多快?  另一种气候威胁:笼罩在阴影与潮湿中的地球
另一种气候威胁:笼罩在阴影与潮湿中的地球  撒哈拉的年龄:从绿色草原到石头沙漠
撒哈拉的年龄:从绿色草原到石头沙漠  1800年至2100年的世界人口
1800年至2100年的世界人口  石油:可再生能源时代巨人的陨落
石油:可再生能源时代巨人的陨落  堪察加拟石蟹:海洋巨人
堪察加拟石蟹:海洋巨人  一个社会的崩溃
一个社会的崩溃  全球生态足迹的轻微进展
全球生态足迹的轻微进展  南大西洋异常区
南大西洋异常区  土壤下沉现象称为地面沉降
土壤下沉现象称为地面沉降  为什么地球经常多云?
为什么地球经常多云?  地球的新云层
地球的新云层 Desertec:欧洲与非洲的太阳能梦想
Desertec:欧洲与非洲的太阳能梦想