
因为进化会以相同的解决方案应对相同的限制条件:当生物生活在相似环境中或面临类似挑战时,自然选择会驱使它们形成相近的形态、功能或行为,即便它们没有最近的共同祖先。这就是趋同演化的原理,它解释了为何海豚与鱼龙拥有相似的水动力流线体型,为何章鱼的眼睛与脊椎动物相似,又为何沙漠植物会演化出相同的生存策略。大自然探索着无数路径,但物理与生物学的法则常将解决方案引回相同的架构。
进化趋同是进化中最迷人的现象之一。它表明,来自相距数亿年的遥远谱系的物种,能够发展出惊人相似的特征。这些相似性作为独立进化的结果呈现,引发了一个问题:所引用的机制是否真的足以解释如此接近的解决方案。
快速游动以躲避捕食者、在深海中感知光线、征服天空、抵抗温度变化、在低氧环境中优化气体交换、承受极端压力、在资源匮乏时节省能量、探测化学信号、或伪装以避免被捕食:这些都是环境施加的约束。
根据理论,缺乏计划、目的或设计的进化机制,一代又一代地探索并选择最有效的解决方案。 但当相距甚远的谱系似乎一次又一次地得出相同答案时,问题便浮现:大自然真的在重新发明,还是遵循着比我们想象中更明显的路径? 仿佛进化被推动着,几乎违背其意愿,去重复某些发明。
这一现象似乎挑战了进化完全依赖偶然与随机的观点。它不再仅仅是巧合,而是一种反复出现的趋势:自然选择如同一位严苛的工程师,推动物种走向有限的有效解决方案。仿佛物质与能量的法则在幕后强加了某些可能的生命形态。用于飞翔的翅膀、用于视物的眼睛、用于制服猎物的毒液……这些创新在毫无关联的谱系中反复出现,以至于暗示着自然或许遵循着一本我们仅能窥见一角的说明书。
古生物学家斯蒂芬·杰伊·古尔德(1941–2002)认为进化在很大程度上是一个不可预测且充满偶然性的过程,但他也承认趋同现象引发了一个令人困扰的问题:如果某些生命形态不仅是偶然的产物,更是深层、或许不可避免的约束条件的表达呢?在他的著作《奇妙的生命》中,他想象“重放生命之带”,并思考:如果进化历史重新开始,同样的生物——或者至少是应对生存挑战的相同解决方案——会再次出现吗?趋同似乎暗示着重复……但也让人对进化中究竟什么是真正的随机性存疑。
这一观点开启了一个令人目眩的视角:生命的多样性,尽管看似繁盛,或许比我们想象的更符合某种可预测的框架。物理世界的约束因此充当了过滤器,只允许某些创新涌现——而这些创新,奇妙地,最终出现在起源相距甚远的物种中。这或许是一堂谦逊的课,但也邀请我们去思考:是否存在更深层、尚未被充分理解的规则,在引导着自然的创造力。
| 假设的趋同特征 | 上述独立团体 | 自适应功能 | 分化时代 | 例子与特殊性 |
|---|---|---|---|---|
| 单镜头眼 | 脊椎动物 / 头足类动物 | 图像形成 | 5.5亿年前 | 人类(盲点)/ 乌贼(无盲点)——尽管共享遗传基因,却存在惊人相似性 |
| 主动飞行(翅膀) | 昆虫 / 翼龙 / 鸟类 / 蝙蝠 | 空中移动 | 3.5亿年前 | 四种不同的体型,但空气动力学限制似乎要求采用相似的解决方案。 |
| 蟹化(螃蟹形态) | 真蟹 / 假蟹(异尾类) | 保护,侧向行走 | 2亿年 | “蟹化”:远缘谱系在相似的限制下,可能演化出相同的外形。 |
| 回声定位 | 蝙蝠 / 海豚 / 金丝燕 | 在黑暗中导航与狩猎 | 80 Ma | 相似的声学机制,但有时却有着惊人趋同的遗传基础 |
| 伪装(同色性) | 昆虫 / 鸟类 / 哺乳动物 | 隐藏 | 3亿年前 | 竹节虫、兰花螳螂、北极野兔——策略相似,但生化途径并非总是独立。 |
| 麻痹性毒液 | 蛇类/慢缺肢蜥/蜘蛛/蝎子/青蛙 | 固定猎物或防御 | 2亿年前 | 类似的毒素,有时基于共同祖先中已存在的基因家族。 |
| 生物发光 | 昆虫 / 深海鱼 / 细菌 / 真菌 / 鱿鱼 | 吸引、诱惑、伪装 | 5亿年前 | 荧光素与荧光素酶:相同的化学反应,但其真正独立的起源仍存争议 |
| 极端嗜性(抗辐射性) | 古菌 / 细菌 / 水熊虫 | 极端电阻 | 2.7 Ga | 类似的DNA修复机制,可能源自非常古老的分子模块 |
在所有进化趋同的例子中,眼睛无疑是最引人注目、或许也是最令人费解的。生物学家越是探究其起源,就越发现简单明了的趋同概念变得复杂起来。
章鱼和人类的共同祖先是一种原始的双侧对称蠕虫,生活在约5.5亿至6亿年前(晚前寒武纪)。这种动物没有具备晶状体、视网膜和瞳孔的结构化眼睛,但并非完全没有视觉工具:它已经拥有原始的光感受器、Pax6基因(眼部发育的真正指挥者),以及整个动物谱系共有的祖先视蛋白。
这正是趋同概念变得不那么明确的地方。当沃尔特·雅各布·格林(1939-2014)在1983年证明,将小鼠的Pax6基因注入果蝇体内可以触发其触角上形成眼睛时,他揭示了眼睛背后的遗传程序在相隔数亿年的动物中是共通的。章鱼和人类或许并非从零开始“重新发明”了眼睛:它们似乎各自以不同的方式重新激活了一套共同的祖先分子工具箱,从而形成了惊人相似的结构。
返祖现象是指现代生物体中重新出现数百万年前消失的古老特征。这一现象表明,某些特征并非由进化真正"重新创造":它们只是从基因组中仍存在但处于沉默状态的祖先遗传程序中被重新激活。鸡长出牙齿、鲸鱼重新出现腿、蛇显现肢芽等现象,都体现了生命体更倾向于重新激活古老结构而非重新创造它们的能力。
因此,返祖现象使进化趋同的解释复杂化:某些被视作独立发明的特征,可能仅仅是预先存在的遗传模块的重新激活。换言之,部分"趋同"并非环境约束催生的新解决方案,而是生命深层历史中刻写的古老状态的可能回归。
查尔斯·达尔文(1809–1882)于1859年提出的理论框架基于三大原则:共同祖先的遗传变异、个体间的可遗传差异,以及作为筛选机制的自然选择。如今,这三大支柱已得到遗传学、古生物学、分子系统发育学及直接观测的证实。科学界中任何严谨的研究者对此均无异议。
达尔文把握住了核心要点,尽管他提出的机制已不足以解释一切。21世纪的生物学并未否定他的成果:而是拓展、完善并丰富了它,有时甚至超越了曾经的想象。
进化趋同展示了环境和物理法则在多大程度上能够引导进化走向相似的解决方案,而非完全听凭偶然。因此,它开启了这样一种可能性——但并非确定性——如果类似的约束条件存在,外星智慧形式也可能在其他地方出现。眼睛、翅膀或类似的社会结构便可能随之出现……或暗示生命有时会遵循比我们想象的更为明确的路径。
然而,这种理论上的可能性不应掩盖一个更深层的现实:进化轨迹在很大程度上仍受制于独特且不可预测的序列。
然而,尽管存在这些令人不安的相似之处,地球生命仍是极其偶然历史的产物,由一系列不可能重复的意外事件塑造而成。没有任何迹象表明这条充满断裂、意外与迂回的道路能在别处重现;相反,一切证据都指向其独特性——它专属地球及其独一无二的历史。
进化趋同指的是没有近期共同祖先的物种独立出现相似特征的现象。这些相似性源于相似的环境压力,促使进化走向类似的解决方案。
不。趋同并非预设的计划:它源于物理、生物和生态的约束,这些约束限制了可行解决方案的数量。进化仍然是偶然的,但并非毫无边界。
因为某些形态在解决类似问题时特别高效:快速游泳、飞行、黑暗中视物、抵抗压力等。环境如同过滤器,筛选出最有效的解决方案。
这表明了一定程度的可预测性:在相似的约束条件下,生物体倾向于进化出相似的结构。然而,生命的多样性表明,许多演化路径仍然是可能的。
鸟类、蝙蝠和昆虫的翅膀;头足类动物和脊椎动物的眼睛;鱼类、海豚和鱼龙的流线型身体;以及蚂蚁和白蚁的社会策略。
是的。多个谱系独立发展出了高级认知能力:头足类动物、鸦科鸟类、灵长类动物以及海洋哺乳动物。这表明,当生态条件有利时,智能便可能涌现。
它提供了一个启发式框架:如果某些解决方案具有普遍效率,那么在相似约束条件下,它们可能在其他星球上再次出现。这为天体生物学中的假设提供了指导。
恰恰相反。这是自然的结果:自然选择探索的是一个受物理、化学和生物学规律限制的可能性空间。趋同现象恰恰体现了这些约束的强大力量。
 最小作用量原理:大自然为何总是选择最经济的路径?
最小作用量原理:大自然为何总是选择最经济的路径?  演化趋同:偶然、必然……还是幻象?
演化趋同:偶然、必然……还是幻象?  五大灭绝事件:这些全球性灾难揭示了什么?
五大灭绝事件:这些全球性灾难揭示了什么?  冰冻圈:当冰之巨人仍在保护我们
冰冻圈:当冰之巨人仍在保护我们 氮循环:从空气到生命
氮循环:从空气到生命  急流:当大气之河决堤
急流:当大气之河决堤  碳循环:海洋、森林与地下土壤的和谐互动
碳循环:海洋、森林与地下土壤的和谐互动  水悖论:没有它,就没有生命
水悖论:没有它,就没有生命 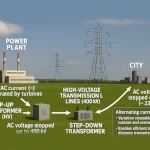 为什么插座里是220V交流电?一个物理学问题
为什么插座里是220V交流电?一个物理学问题  最后一次冰川冲击:新仙女木期及其对史前社会的影响
最后一次冰川冲击:新仙女木期及其对史前社会的影响  536年:没有夏天的一年与晚古小冰期的开始
536年:没有夏天的一年与晚古小冰期的开始  最小细胞:不可避免的涌现
最小细胞:不可避免的涌现 

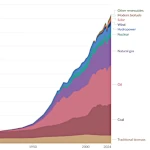 全球能源消费演变:从蒸汽机到数字服务器
全球能源消费演变:从蒸汽机到数字服务器  人口转型:增长还是衰退?
人口转型:增长还是衰退?  阿加西湖:改变气候的洪水
阿加西湖:改变气候的洪水  醉林:理解永久冻土融化的现象
醉林:理解永久冻土融化的现象  空气中有电!
空气中有电!  为什么二氧化碳不会掉落到地面?
为什么二氧化碳不会掉落到地面?  厄尔尼诺与拉尼娜:塑造全球气候的太平洋两股呼吸
厄尔尼诺与拉尼娜:塑造全球气候的太平洋两股呼吸  濒危物种:灭绝动力学
濒危物种:灭绝动力学  酸雨的有毒遗产
酸雨的有毒遗产  全球变暖数据:科学数据告诉我们什么
全球变暖数据:科学数据告诉我们什么  什么是可持续发展?
什么是可持续发展?  咸海:给人类的一课
咸海:给人类的一课  三峡大坝:对生态系统的影响
三峡大坝:对生态系统的影响  有记录以来最热的年份
有记录以来最热的年份  未来全球变暖的幅度
未来全球变暖的幅度  危机中的地球:崩溃还是复兴?
危机中的地球:崩溃还是复兴?  冰层告急:北极海冰不可阻挡的衰退
冰层告急:北极海冰不可阻挡的衰退  地球的水库:从海洋到地下水
地球的水库:从海洋到地下水  海平面正在上升,但速度有多快?
海平面正在上升,但速度有多快?  另一种气候威胁:笼罩在阴影与潮湿中的地球
另一种气候威胁:笼罩在阴影与潮湿中的地球  撒哈拉的年龄:从绿色草原到石头沙漠
撒哈拉的年龄:从绿色草原到石头沙漠  1800年至2100年的世界人口
1800年至2100年的世界人口  石油:可再生能源时代巨人的陨落
石油:可再生能源时代巨人的陨落  堪察加拟石蟹:海洋巨人
堪察加拟石蟹:海洋巨人  一个社会的崩溃
一个社会的崩溃  全球生态足迹的轻微进展
全球生态足迹的轻微进展  南大西洋异常区
南大西洋异常区  土壤下沉现象称为地面沉降
土壤下沉现象称为地面沉降  为什么地球经常多云?
为什么地球经常多云?  地球的新云层
地球的新云层 Desertec:欧洲与非洲的太阳能梦想
Desertec:欧洲与非洲的太阳能梦想